
Генетика и ДНК. Роберт Картер, доктор философии (морская биология) [Университет Майами]
Роберт Картер значительную часть своей докторантуры изучал генетику белков, вырабатываемых медузами и кораллами, – зелёного флуоресцентного белка (ЗФБ) и других белков этого семейства. С тех пор он сфокусировал свои усилия на изучении генома человека. Результатом первого его исследования была публикация консенсусной последовательности самого первого митохондриального генома человека – фактически, Евы. В настоящее время он занимается изучением некоторых аспектов генетики человека, в том числе исследованием типологии разнообразия и мутаций, и их взаимосвязи с библейской моделью генетики, следующей из Бытия.
После некоторых несерьёзных попыток в молодости примирить Бытие и эволюцию он на первом курсе университета принял эволюцию как «факт». Позже ему пришлось пересмотреть свои взгляды, отбросив эволюцию в пользу библейского сотворения, но только лишь в первый год докторантуры он полностью убедился в обоснованности позиции младоземельных креационистов.
Как креационист, глубоко разбирающийся в генетике, а также как бывший эволюционист (пусть и в течение недолгого времени) он написал главу, рассматривающую вторую ахиллесову пяту эволюции – генетику.
В главе 1 мы увидели, что естественный отбор не был детищем ума Дарвина. Мы также увидели, что его недостаточно для обоснования главной идеи Дарвина – что все виды связаны общим происхождением. В этой главе мы рассмотрим кое-что, о чём Дарвин не имел представления, – генетику. Несмотря на его незнание генетики, сейчас невозможно говорить об эволюции путём естественного отбора, не касаясь вопроса потенциальных изменений в наследственном материале жизни. Ответы на вопросы типа
«Как гены передаются из поколения в поколение?» и «Какая частота мутаций допустима для выживания популяции в течение длительного времени?» являются ключевыми в понимании этой ахиллесовой пяты эволюции.
Краткая история генетики
Философы тысячелетиями спорили о способах, с помощью которых организмы передают свои признаки потомству. На протяжении большей части человеческой истории мы не знали, что приводит к вариациям внутри видов или как эта вариация передаётся из поколения в поколение. До конца 19-го столетия большинство людей, похоже, придерживались идеи смешанной наследственности. То есть, что потомство является комбинацией или усреднением своих родителей. Это в некотором роде имело смысл, поскольку легко проследить признаки обоих родителей во многих детях, но не означает ли это, что все признаки должны были усредниться со временем? Люди знали об этой проблеме, но на тот момент не было предложено какой-то лучшей идеи.
Поскольку Чарльз Дарвин жил в то время, он не имел никакого представления о генетике. То есть его идеи эволюции были развиты в некотором вакууме, полном умозрительных гипотез. Будучи неудовлетворённым превалирующими идеями наследственности, Дарвин изобрёл свою собственную теорию, называемую пангенезис.92 Он представлял себе «корпускулы», которые производятся различными органами тела в ответ на раздражения со стороны окружающей среды. Эти корпускулы затем якобы должны отправиться в половые железы и храниться там, пока не передадутся следующему поколению. Однако, не было никакого физического или экспериментального обоснования для такого взгляда. Также он напрямую связан со старой теорией под названием ламаркизм, или наследственность по Ламарку. Жан Батист Ламарк (1744–1829) учил, что организмы отвечают на окружающую среду, становясь более крепкими, более высокими, более дальнозоркими, или, например, способными выдержать избыточное тепло. Затем организм передаёт вновь приобретённые признаки своему потомству, которое начинает свою жизнь, будучи немного сильнее, выше и т.д., чем были изначально родители. Фраза, которая ассоциируется с такими взглядами, – «наследование приобретённых признаков». Дарвин, конечно, примкнул к этому взгляду,93 но он ошибся.
Самое странное, что эта дарвиновская теория пангенезиса была опубликована непосредственно после открытия отцом современной генетики, Грегором Менделем, законов генетической наследственности (впервые опубликованных в 1866 г.94). Работая в монастырском саду, Мендель понял, что многие признаки проявляются в дискретных вариантах (белые или фиолетовые цветки, жёлтые или зелёные семена и т.д.), каждый из которых
называется аллелем. Он также определил, что каждый индивидуум несёт два аллеля для каждого признака (т.е. белый + белый, белый + фиолетовый или фиолетовый + фиолетовый) и что каждый родитель отдаёт только один из аллелей своему потомку. Потомок, таким образом, является не средним (смесью) своих родителей, а комбинацией дискретных признаков, переданных родителями детям. Мендель был монахом, но он не был на «вы» с наукой, получив университетское образование в области физики и философии. Во времена Менделя «церковь» была тем местом, где проводилась значительная часть научной работы, о чём свидетельствует теологическое и научное образование Дарвина, полученное им в Кембридже,95 когда он планировал стать (но так никогда и не стал) священником.
Дарвин имел все те же инструменты, которые были у Менделя, в его распоряжении было больше времени, и, возможно, он имел лучшие финансовые возможности для своих исследований, однако же он не открыл ничего подобного современной теории генетики. Дарвин сделал кропотливые описания генетической изменчивости и закономерностей наследования многих видов растений и животных, в одном случае даже отметив знаменитое менделевское соотношение 3:196 доминантного признака к рецессивному при скрещивании двух гибридов.97 Дарвин написал ряд эссе и статей по этим вопросам.98 Почему же тогда он не открыл генетику? Невозможно знать наверняка, но Дарвин хотел, чтобы виды имели неограниченную способность мутировать, поскольку он был приверженцем непрерывного распределения видов, с бесчисленными переходными звеньями между ними, идущими бесконечно в прошлое. Это повлекло за собой (или вписывалось в) поиски непрерывного распределения признаков, результатом чего стало возникновение теории пангенезиса и его обращение к ламаркизму. Но гены не кодируют непрерывные распределения. Они кодируют дискретные признаки. Хотя действительно многие признаки определяются совместным влиянием многих различных генов (создавая впечатление непрерывности), это всего лишь делает связь между окружающей средой и генетикой намного более сложной. Дарвин смотрел в неверном направлении. Мендель был экспериментатором и пришёл к правильному ответу. Дарвин был теоретиком и мог упустить правильный ответ исключительно из философских предпосылок.99
Но это не было ошибкой одного лишь Дарвина, поскольку большинство людей того времени также не увидели её. Лишь несколько десятилетий спустя идеи Менделя наберут популярность. Но в течение всей своей жизни Дарвин не имел представления о фундаментальных процессах, на основе которых должна была бы строиться теория эволюции. Однако оказывается, что законы генетики недружелюбны к теории эволюции.
Современная генетика
C начала 20 столетия генетические исследования начали набирать обороты. Было открыто, что совершенно новые признаки, названные мутациями, могут внезапно появляться у индивидуума, и работы Менделя оказались востребованы. Потребовалось приложить немало усилий, но к 1920-м годам мутации, естественный отбор и генетика были объединены в так назы- ваемую синтетическую теорию эволюции. Смешанная наследственность, пангенезис и ламаркизм были отброшены в пользу нового понимания, которое не сильно отличается от того, во что верят большинство эволюционистов сегодня, около 150 лет спустя.
Область генетики медленно развивалась в первой половине столетия, но в 1950-х годах произошёл взрыв. В 1952 г. знаменитый эксперимент Херши–Чейз доказал, что именно ДНК является носителем наследственности (факты, указывающие на это, были и прежде, однако многие люди всё ещё считали, что эту функцию выполняют белки). В 1953 г. Уотсон и Крик открыли, что ДНК имеет форму двойной спирали. Эксперимент Мезельсона–Сталя в 1958 г. показал, что две цепочки ДНК рассоединяются и копируются независимо друг от друга. Вскоре после этого был расшифрован белковый код, и мы узнали, что три буквы ДНК кодируют одну аминокислоту в белке.
Фундамент современной генетики был заложен. Секвенирование генов было разработано в последующие десятилетия. Затем оно было автоматизировано. Смена тысячелетий возвестила нам новый мир, так как в 2003 году проект «Геном человека» опубликовал транскрипт около 93% линейных цепочек ДНК из ядра клетки человека. Дерзкий по размаху, масштабам и финансированию, этот проект по определению последовательности 3 млрд букв ДНК стоил американским налогоплательщикам около 3 млрд долларов США. Это монументальное (и дорогое!) достижение всё изменило. Сегодня, с быстрым развитием технологий, есть компании, которые могут легко секвенировать эквивалент нескольких геномов человека ежедневно, и они могут сделать это всего за несколько тысяч долларов каждый. И даже это будет прошлым днём всего через несколько лет. Технологические достижения позволяют совершать новые и новые открытия быстрыми темпами, и многим из наших традиционных представлений брошен вызов.
Данные накапливаются быстро. Пару десятилетий назад нельзя было и помыслить об ответах на вопросы, которые имеются сегодня. Возможно, впервые в истории, у нас появилась обширная научная область, в которой больше данных, чем теории! Добро пожаловать в мир современной генетики.
ДНК как носитель информации
Всё живое зависит в своём выживании и воспроизводстве от специфичной информации (своего рода набора инструкций). Большая часть этой информации содержится в ДНК-молекуле, которая является непревзойдённым носителем информации. В совершенно незначительном количестве ДНК можно записать невероятно большое количество инструкций. Возьмём лист бумаги с напечатанным текстом. Каждая буква на этом листе примерно 3 или 4 мм высотой, и обычная страница может содержать несколько тысяч букв. Если напечатать геном человека в текстовом виде в книгах размером с Библию (может отличаться в зависимости от перевода, например, Библия на английском содержит около 3,5 млн букв), то нужно было бы около 850 экземпляров, чтобы поместить его весь. И поскольку вы наследуете один экземпляр от своей матери, а второй – от своего отца, вы содержите две копии генома в каждой клетке вашего тела (за исключением красных кровяных клеток, которые не имеют ядра). Буквы ДНК, однако, размером всего лишь в несколько миллиардных долей метра. При таких размерах все три миллиарда букв могут легко поместиться внутри чего-то гораздо меньшего, чем точка в конце этого предложения. Внутри клетки они связываются вместе в цепочки (хромосомы), так что, если выстроить их одну за другой, полученная цепочка будет составлять около 2-х метров в длину. Эти длинные, хрупкие, липкие нити упакованы в ядре всего 6–10 мкм (миллионных долей метра) в диаметре. Весь набор команд для создания и функционирования человека хранится в чём-то столь малом, что невозможно увидеть без микроскопа!
Но эти цепочки не очень стабильны. Однонитевые цепочки ДНК подвержены распаду. В ядре клетки каждая нить соединена с комплементарной нитью, и они закручиваются относительно друг друга, образуя классическую форму двойной спирали. Участок ДНК является фактически двумя отдельными, но комплементарными молекулами, которые «склеены» друг с другом с помощью электромагнитного взаимодействия. Три миллиарда букв ДНК генома человека находятся в 23 отдельных хромосомах, каждая из которых имеет от примерно 50 до приблизительно 250 миллионов букв в длину. Но даже в таком виде ДНК не является очень стабильной. Она всё ещё очень чувствительна к деградации, и, по оценкам, до одного миллиона «повреждений» ДНК (например, разрыв цепочки, повреждения, вызванные радиацией, или разрушение отдельных букв-оснований путём реагирования с кислородом) происходит в обычной клетке ежедневно100 (а наше тело состоит из около 100 триллионов клеток!). Представьте себе усилия, которые клетка ежедневно должна тратить, чтобы поддерживать свою копию ДНК в порядке. Теперь умножьте эти усилия на количество клеток. 1000000 ´ 100000000000000 – именно столько исправлений каждый день необходимо, чтобы геном продолжал работать в нашем теле.
Хрупкость ДНК является одной из многих ахиллесовых пят эволюционной генетики, но при этом одной из самых важных. Чтобы ДНК могла быть полезной, необходим огромный вспомогательный набор ремонтирующих ферментов для поддержания её целостности. Повреждения ДНК могут быть различными, и для каждого рода повреждений существуют специализированные комплексы ферментов. Но ещё большей трудностью для эволюционной модели является то, что сами эти ферменты также закодированы в ДНК, хотя без них ДНК не может поддерживаться в клетке. Это совершенная проблема курицы и яйца! Эти ферменты также чувствительны к изменениям. Мутации в участках ДНК, которые кодируют ремонтирующие и копирующие ферменты, часто являются катастрофическими. Как тогда они могли произойти путём процесса мутаций и отбора с течением времени? Без них жизнь не может существовать, однако жизнь должна была начинаться без них и должна была начать использовать ДНК для хранения информации прежде, чем эволюционировал набор инструментов для работы с ДНК. Поскольку это будет более детально рассмотрено в следующей главе, посвящённой происхождению жизни, достаточно будет сказать, что ДНК – последний выбор носителя информации, которого можно было бы ожидать от первичной жизни.
ДНК и теория информации
Порядок, в котором нуклеотиды выстроены в молекуле ДНК, обладает всеми характеристиками сообщения, перегруженного информацией.101,102 Но в реальной жизни не бывает сообщения без отправителя. Сложнейшей загадкой жизни является не сложность молекулы, на которой основана жизнь (хотя это и огромная загадка). И не сложная организация компонентов, из которых построены живые организмы (хотя это ещё одна огромная загадка). Сложнейшей загадкой жизни является происхождение информации, на которой основана жизнь. Это уже затрагивает вопрос происхождения жизни, что не является предметом этой главы. И что нас больше всего заботит – это поддержание целостности информации в геноме в течение жизни индивида и от поколения к поколению.
Это является ещё одной ахиллесовой пятой эволюционной генетики, так как она не работает без почти идеальной системы поддержания целостности информации. И даже несмотря на потрясающие ремонтные системы ДНК в теле человека, мы всё равно накапливаем около 100 новых мутаций на каждого человека с каждым новым поколением.103 По оценкам, при каждом делении клеток происходит от одной до трёх мутаций.104 Это неплохая частота ошибок для маленьких молекулярных машин, отвечающих за быстрое копирование ДНК непосредственно перед клеточным делением. Но посмотрите на это в контексте количества клеточных делений, необходимых для формирования человека: в настоящее время несёте все возможные мутации, неоднократно, и их количество только увеличивается со временем. Из-за этого накопления ошибок со временем, по оценкам, 60-летний человек имеет 40000 мутаций в каждой из клеток, выстилающих кишечный тракт.105 На самом деле, за исключением несчастных случаев и болезней, это неизбежное накопление ошибок приведёт к смерти каждого из нас.
К счастью, Создатель (Бог) разработал блестящий механизм защиты человечества от последствий этих мутаций. После оплодотворения яйцеклетки зигота проходит через ограниченное количество клеточных делений прежде, чем сформируются клетки для следующего поколения. В ребёнке-девочке происходит всего 23 клеточных деления до завершения формирования яичников. Яйцеклетки, находящиеся в яичнике, разовьются и будут защищены до овуляции, иногда 40 или более лет, без дальнейшего деления. Мало кто знает, что в организме женщины, вынашивающей девочку, одновременно присутствуют три поколения: мама, девочка, и яйцеклетки в лишь недавно сформированных яичниках ребёнка. У мужчин, однако, всё немного отличается. Мужчина проходит через тридцать делений в утробе матери, прежде чем его репродуктивные клетки будут готовы, но они начинают быстро делиться при наступлении половой зрелости и будут продолжать деление до самой смерти. Это означает, что отец передаёт больше мутаций ребёнку, зачатому в старости, чем ребёнку, зачатому в молодости.106
Ограниченные исследования мутаций, передаваемых от родителей к детям, дали, тем не менее, некоторые удивительные результаты. Оказывается, что число мутаций, получаемых от каждого из родителей, может быть весьма различным, иногда больше передаётся от отца, а иногда больше и от матери.107 Это вызов стандартным моделям генетической истории человека, которые основаны на молекулярных часах, предполагающих равные частоты мутаций в ходе времени и по географическому ареалу человечества. Мы обсудим последствия этого более подробно ниже.
Хотя большинство этих мутаций не являются катастрофичными (иначе мы все бы уже вымерли), они всё равно плохие. Генетики называют их слегка вредными мутациями, и их накопление в геноме человека – это серьёзный вызов эволюционной теории.108 Эти плохие мутации накапливаются со скоростью, превышающей ту, с которой естественный отбор мог бы теоретически их удалить. Вообще, лишь самые худшие мутации попадают в поле зрения естественного отбора (глава 1). Поэтому эволюция движется в обратном направлении, будучи неспособной предотвратить медленную деградацию информации, необходимой для жизни. Если это правда, то как же тогда информация могла появиться изначально? И как удалось видам выжить и не вымереть за все эти миллионы лет?
«Мусорная» ДНК является (большей частью) функциональной!
Десятилетиями мы слушали старую «пластинку» про то, что «в геноме человека лишь от 2 до 3 процентов функциональны. Остальная часть – бесполезная, мусорная ДНК – мусор, доставшийся нам как наше эволюционное наследие». Хотя сказанное до сих пор является распространённым убеждением, недавние открытия показали, что оно неверно. Почему же тогда мы слышали об этом так часто в течение длительного времени? Потому что эволюционной биологии необходима мусорная ДНК, чтобы решить серьёзную математическую проблему.
В конце 1950-х знаменитый популяционный генетик Джон Холдейн показал, что естественный отбор не был способен выбрать миллионы полезных мутаций, даже в течение предполагаемой человеческой эволюционной истории. Даже несмотря на несколько упрощающих допущений в пользу эволюционной теории,109 всего несколько сотен полезных мутаций могли бы быть выбраны со времени нашего общего предка с шимпанзе.110 Эта проблема была названа дилеммой Холдейна, и, несмотря на многочисленные заверения в обратном, она так и не была решена.111 Вместо решения был предложен плод эволюционного воображения. В конце 1960-х годов Кимура развил идею нейтральной эволюции.112 Он рассуждал, что если бы большая часть ДНК в клетке была нефункциональной, то она могла бы свободно мутировать в течение длительного времени. Таким образом, стоимость поддержания нефункциональных частей генома для популяции была бы нулевой («стоимость» измеряется количеством дополнительного потомства, которое должно рождаться в популяции и которое естественный отбор мог бы уничтожить, чтобы устранять плохие мутации и поддерживать приспособленность в течение долгого времени,113 конечно, если предположить, что естественный отбор может «видеть» плохие мутации, которые необходимо устранять).
Изобретение термина «мусорная ДНК» примерно четырьмя годами позже приписывают Сузуму Оно.114 Идея мусорной ДНК чрезвычайно важна для эволюционных математиков. Что бы произошло, если бы оказалось, что она неверна? Что произойдёт, если вместо того, чтобы на 97% быть мусорным, геном окажется на 97% функциональным?
Современные технологии в настоящее время убили понятие мусорной ДНК. Это произошло после завершения проекта «Геном человека». Впрочем, и раньше было ясно, что оно ошибочно, но, похоже, сказывалось упорное нежелание отказаться от теории мусорной ДНК. Есть много причин полагать, что бо́льшая часть ДНК является функциональной. Например, были установлены функции многих ретротранспозонов,115 которые когда-то посчитали кусочками вирусов, которые интегрировались в наш геном в течение миллионов лет.
Кроме того, были установлены функции для большинства обширных участков не кодирующей белок ДНК, которая находится между генами. Оказывается, большая часть генома является активной. Проект ENCODE [прим. пер.: The Encyclopedia of DNA Elements – «Энциклопедия элементов ДНК»] – это многолетнее исследование стоимостью несколько миллионов долларов при участии ряда университетов, целью которого было определить, какая часть человеческого генома транскрибируется (копируется в РНК, что является показателем наличия функции). Был проанализирован только 1% генома, но в него включили как кодирующие белок, так и «мусорные» участки ДНК. Этот проект показал, что в среднем каждая отдельная буква генома используется в шести различных транскриптах РНК.116 Это отнюдь не означает, что всё превращается в белок. Это также не означает, что всё имеет конкретную функцию или даже что буквы часто используются. Это означает то, что почти каждая буква делает что-то. Так как «форма следует за функцией» является общим правилом биологии, тот факт, что эти участки являются активными, явно свидетельствует о том, что у них есть функции. Почему бы ещё клетки позволяли такой большой объём транскрипции? Клетка тратит значительную часть своих ресурсов на синтез РНК, не кодирующей белок. То есть клетка бы сильно выиграла, отключив такую растрату энергии. Естественный отбор в течение миллионов лет покончил бы с этим паразитическим процессингом РНК. Он не сделал так потому, что этот процессинг необходим для функционирования клетки. Фактически, геном теперь можно рассматривать как РНК-компьютер (смотрите ниже).
Общая нехватка мусорной ДНК – ещё одна ахиллесова пята эволюционной генетики, потому что без неё эволюционная математика не работает, а у ДНК, не кодирующей белок, продолжают обнаруживаться всё больше и больше функций. На самом деле, ситуация, когда «мусорная» ДНК, похоже, более активна, чем «гены», переворачивает с ног на голову старую идею, что мы – белковые организмы. По словам эволюционного биолога Джона Маттика:
Непризнание всех последствий этого, особенно возможности того, что промежуточные некодирующие последовательности могут передавать информацию параллельно в виде молекул РНК, вполне может оказаться одной из самых больших ошибок в истории молекулярной биологии.117
Ультра сложная обработка генов
Студентов-биологов прошлого всегда учили гипотезе «один ген – один фермент». На основании потрясающих открытий, совершённых в течение 20 века, очевидным казался вывод, что «ген» – это участок ДНК, кодирующий определённый белок. «Ген» имел определённые начальную и конечную точки, секции, кодировавшие белок (экзоны), иногда несколько промежуточных последовательностей (интронов), которые необходимо отрезать от первичного РНК-транскрипта перед тем, как «ген» будет транслироваться в белок, и регуляторные области, расположенные по ходу и против хода транскрипции, к которым могут присоединяться различные эпигенетические факторы, контролирующие экспрессию «гена». Было несложно двигаться вдоль «гена» и видеть эти места. Можно было даже сходу транслировать генетический код, содержащийся в последовательности на цепочке информационной РНК, в белок, зная трёхбуквенные коды для каждой аминокислоты (что не очень сложно). Однако существует проблема с гипотезой «один ген – один фермент»: как и гипотеза пангенезиса Дарвина и идея мусорной ДНК, она также неверна.
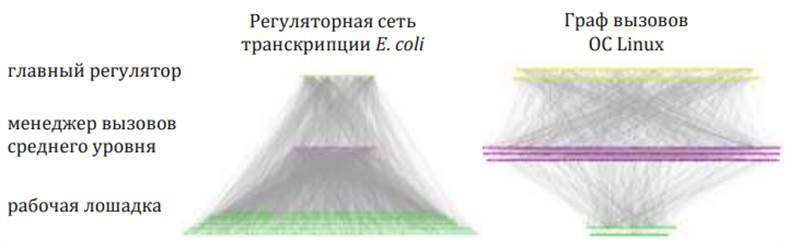
Сравнение программной архитектуры бактерии E. coli и графа вызовов операционной системы Linux показывает интересные сходства и поразительные различия между двумя системами. Из Yan и др., 2010 г.118
Благодаря результатам проекта ENCODE мир смог заглянуть внутрь самой сложной компьютерной операционной системы в известной Вселенной – внутрь человеческого генома. Но это не белковый компьютер. На самом деле, геном – это скорее рибонуклеиновый суперкомпьютер, который на выходе выдаёт белок. Подобно вашему компьютеру, который имеет жёсткий диск, перезаписываемую память (RAM) и экран для вывода, геномный компьютер имеет ДНК для хранения информации, РНК для сравнения и обработки информации и расчёта, а также белки как средство вывода.
Были предприняты попытки сравнить процессы управления в геноме и в созданных человеком компьютерных системах.119 Параллели интересны, но различия просто поразительны. Например, по сравнению с компьютерной операционной системой Linux бактериальный геном кишечной палочки (E. coli) имеет меньше высокоуровневых регуляторов, которые управляют меньшим количеством контроллеров среднего уровня, которые в свою очередь управляют намного большим количеством низкоуровневых средств вывода. Создаётся впечатление, что её геном оптимизирован, чтобы делать то, что он делает, настолько эффективно, насколько это возможно. Это намного больше похоже не на Linux, а на военные компьютеры, которые, как правило, имеют очень короткие программы с минимальными инструкциями. Это верно, потому что они предназначены выполнить как можно эффективнее ограниченный набор действий, а не много разных действий одновременно (как, например, одновременное использование графики, игр, музыки и текстового редактора, что позволяет Linux). Тем не менее, здесь одна загадка: потребовалось множество человеко-часов, чтобы спроектировать компьютеры, которые стоят на борту бомбардировщиков В-52, и всё же одна ошибка в любой программе любой подсистемы может вызвать катастрофический отказ системы, которую она контролирует. Геном человека является гораздо более сложным и может выдержать тысячи ошибок без сбоев. Он контролирует больше вещей одновременно и лучше разработан!
Когда секвенирование генома человека было завершено, исследователи были шокированы, что в нём было найдено всего лишь около 23000 «генов».120 К тому времени уже было известно, что количество белков, которые производит человеческое тело, во много раз превышает это число. Как такое возможно? Через несколько лет проект ENCODE вполне определённо указал нам на то, что в геноме человека широко применяется альтернативный сплайсинг.121 Мы узнали, что каждая часть «гена» может быть использована в нескольких различных белках.122 Каким-то образом тело знает, как создавать различные комбинации того, что мы считали отдельными кодирующими белок генами, и соединять их вместе, чтобы создать несколько сотен тысяч уникальных белков. К тому же, разные типы клеток могут создавать различные белки посредством этого сложного процесса. Кроме того, различные белки производятся в разное время, и каким-то образом клетки знают, что производить, когда это производить и при каких условиях.123 Существует что-то, что контролирует этот процесс, и оно не обязательно находится в кодирующей белок части генома.124
В каждом «гене» вложена серия небольших кодов. Каждый длиной всего в несколько букв, но есть несколько кодов в начале и в конце каждого экзона и интрона. Они составляют то, что называется сплайсосомой, то есть частью генома, которая контролирует сложный процесс рекомбинации экзонов, или сплайсинга генов. Сложность сплайсосомы, фактически, сложность всего генома всех эукариот, – это ещё одна ахиллесова пята эволюционной генетики. Геном является слишком сложным, а «мишень» для мутаций – слишком большой, чтобы известные виды смогли выжить в течение миллионов лет,125 не говоря уже о том, чтобы они прежде эволюционировали.
У вас, возможно, возник вопрос: почему в этой главе я поставил слово «ген» в кавычки? Это потому, что у нас больше нет определения этого слова.126 По крайней мере, определение изменилось, и теперь означает нечто совершенно новое для всех организмов, более сложных, чем бактерии. Генетика сделала поворот в сторону сложности, и прежний, простой взгляд стал устаревшим. Отсюда и далее слово «ген» понимайте в классическом смысле: ген – это участок ДНК, который кодирует белок. Единственная проблема с этим определением, что любой отдельный участок ДНК может быть использован в нескольких белках в зависимости от контекста.
Гиперсложность четырёхмерного генома
Давайте перейдём к чему-то ещё более интересному и посмотрим на другой уровень сложности. Когда мы секвенировали геном человека, мы думали, что теперь поймём, как работает геном. Это было наивной ошибкой. То, что мы сделали, было всего лишь определение линейной последовательности нуклеотидов. Это было лишь первое измерение генома, который работает по крайней мере в четырёх измерениях. Что это значит? Подумайте о ДНК. Это строка, а строка, по определению, является одномерной. Когда секвенирование человеческого генома было завершено, казалось, что теорию мусорной ДНК подтвердил тот факт, что гены были разбросаны по всему геному и что нет никакой кластеризации связанных функций. «Ага», – сказали исследователи, – «порядок генов является случайным, продукт случайных изменений с течением времени». Однако это было немного близоруко, ведь они смотрели всего лишь на первое измерение.
Мы только что узнали об альтернативном сплайсинге. Здесь одна часть генома влияет на другую непосредственно или посредством РНК и/или белка. Это является частью второго измерения генома. Для того чтобы нарисовать эти взаимодействия, нужно было бы выписать геном и нарисовать много-много стрелок из одного места в другое. Чтобы сделать это, вам потребуется много листов бумаги, которые имеют два измерения (высоту и ширину).
Второе измерение генома является чрезвычайно сложным и включает в себя факторы специфичности, усилители, репрессоры, активаторы, транскрипционные факторы, сигналы ацетилирования гистонов, сигналы метилирования ДНК, посттранскрипционное регулирование РНК, альтерна тивный сплайсинг и многое другое. Оно играет важную роль в жёсткой координации и регулировании обширной сети событий, которые происходят как в ядре, так и по всей клетке. В этом измерении порядок не особенно важен, поскольку регуляторы генов в любом случае должны передвигаться, чтобы найти свои мишени. То есть нахождение мишени непосредственно рядом с регулятором не обязательно. Именно на следующем уровне всё становится очень интересно.

Первые несколько сотен букв человеческой Y-хромосомы являются хорошим примером первого измерения генома. Это просто строка из A, T, Г и Ц.
Третье измерение генома – это трёхмерная структура ДНК в ядре. На этом уровне гены распределены в ядре не случайным образом, а упорядочены и сгруппированы с учётом конкретных потребностей. Гены, которые используются вместе в серии, могут не находиться друг возле друга на хромосомах, но когда хромосомы сворачи ваются, они часто оказываются рядом друг с другом в трёхмерном пространстве, а также часто группируются вблизи ядерных пор или рядом с центром транскрипции.127
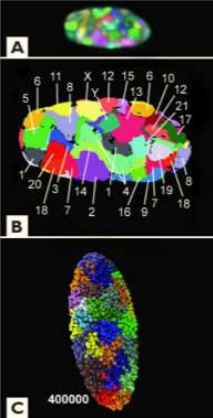
A: изображение, полученное путём окрашивания хромосом фибробластной клетки человека флуоресцентными красителями. B: псевдоцветное представление тех же данных. C: компьютерная модель организации хромосом
То есть что-то удерживает их в нужном месте. Поскольку ДНК хранится в упакованном виде, то её заглублённые участки являются труднодоступными, другие же участки направлены наружу или во внутренние полости.128,129 Часть кода, внедрённая в первом измерении, влияет на трёхмерное сворачивание ДНК, которое в свою очередь влияет на экспрессию генов. Это третье измерение является чрезвычайно важным.130
Четвёртое измерение генома подразумевает изменения в первом, втором и третьем измерениях с течением времени. В ядре хромосомы приобретают определённую конфигурацию, но эта конфигурация изменяется во время развития, так как различным типам клеток нужны разные наборы генов и других генетических инструкций. Конфигурация может измениться за короткий срок, когда клетки реагируют на раздражители и разворачивают фрагменты ДНК для того, чтобы получить доступ к заглублённым генам, только чтобы вновь свернуть этот участок, когда ген больше не нужен.131 Есть также изменения в «мусорной» ДНК. Например, значительная часть активности ретротранспозонов (также известных как прыгающие гены или мобильные генетические элементы) происходит по мере развития мозга, когда участки ДНК нескольких различных классов (L1, Alu и SVA) копируются и прыгают по геному отдельных клеток мозга. Это помогает различным мозговым клеткам дифференцироваться.132 Кроме того, клетки печени, как правило, имеют много дубликатов хромосом. Окончательный геном различных клеток в организме не обязательно такой же, каким был в оплодотворённой яйцеклетке, когда она начала делиться, кроме того, конфигурация генома варьируется от клетки к клетке и с течением времени. Эти примеры – не случайности, а тщательно контролируемая симфония геномных изменений в четырёх измерениях. Я надеюсь, вы впечатлены, так как геном был разработан Великим Архитектором.
Всё это невероятно сложно, но подчёркивает очередную ахиллесову пяту эволюционной генетики. Дарвинизму необходимо, чтобы жизнь была простой. Естественный отбор должен иметь возможность брать небольшие изменения в результате мутаций и выбирать лучшее из стада или группы животных. Когда вид уже существует, возможно, естественный отбор может действовать ограниченным образом (см. предыдущую главу), но может ли этот процесс объяснить, как вид появился изначально? Вряд ли, ведь простой процесс накопления ошибок и отбора не может создать сложную, с перемежением, четырёхмерную систему с удивительной степенью сжатия данных и гибкостью. И когда эта система уже существует, для неё серьёзной угрозой будут дальнейшие случайные изменения посредством мутаций. Это та самая ситуация, в которой мы находимся сегодня. Можно представить небольшие изменения в уже существующей сложной системе. Но использовать эти небольшие изменения в качестве объяснения происхождения самой этой системы, однако, равносильно утверждению, что в создании новейших компьютерных операционных систем не участвовал разум. А геном намного превосходит по сложности и эффективности любую современную операционную систему в мире.
Избыточность кодона
Сегодня созданные человеком компьютеры работают в двоичной системе счисления, потому что транзисторы имеют лишь два состояния и, соответственно, дают нам лишь два символа, с которыми можно работать (0 и 1 – выкл. и вкл.), но математика, основанная на двоичной системе, довольно проста и позволяет строить сложные компьютерные процессоры, работающие в этой системе. Геном же работает в четверичной системе. Вместо 0 и 1 генетический код состоит из четырёх символов (А, Ц, Г и Т). Это не является обязательным, та же информация могла бы быть записана в системе, использующей любое количество символов. Почему же именно четверичная система?
Интересно, что для кодирования одной аминокислоты в белке нужно три буквы в геноме. В геноме эти трёхбуквенные кодоны соединены в группы (экзоны), каждая из которых производит функциональную часть одного или более белков. В белках клеток человеческого тела используются только 20 аминокислот, но существуют 64 возможных кодона (три позиции с четырьмя возможными буквами позволяют 4 ´ 4 ´ 4 = 64 кодона). Это означает, что некоторые аминокислоты, такие как аланин, кодируются различными кодонами (GCA, GCC, GCG и GCT). Другие, такие как триптофан, – всего одним (TGG). Можно заметить, что все четыре кодона аланина начинаются с букв «GC». Это означает, что любая мутация, которая изменит последнюю букву, будет производить ту же аминокислоту, несмотря на то, что кодон изменился. Это называют избыточностью кодона, и она добавляет немного надёжности геному, поскольку по крайней мере некоторые мутации в кодирующих белок областях будут оказывать незначительное влияние. Я сказал «незначительное», потому что транспорт ные РНК для различных кодонов встречаются в клетке с разной частотой. Известны случаи, когда изменение одной буквы, хотя аминокислота и не была изменена, являлось плохой мутацией. В процессе трансляции белка возникала пауза, пока подыскивалась редкая тРНК. Это вызывало неправильное сворачивание белка, то есть деформированный фермент. Более свежие данные показывают, что используемые кодоны могут влиять на трансляцию белка в целом, по крайней мере у бактерий, у которых скорость трансляции зависит от того, какие именно кодоны используются. Бактериальные клетки избегают генных промоторов внутри их генов. Когда они встречаются, трансляция обычно временно замедляется, потому что бактериальная рибосома имеет обыкновение «залипать» на некоторых последовательностях промоутеров.133 Таким образом, наличие альтернативных кодонов позволило Создателю грамотно спроектировать гены с меньшим количеством внутренних противоречий с другими функциями гена.
Кроме всего этого, есть ещё одна очень веская причина для избыточности кодонов и четверичной системы кодов генома. Это не только самый оптимальный способ закодировать 20 аминокислот,134 но и возможность использовать несколько «перекрывающихся кодов».
Перекрывающиеся коды
В геномах всех высших организмов наблюдается значительная степень сжатия данных. Любой конкретный участок каждого из этих геномов может одновременно быть задействован в нескольких процессах. Одна буква ДНК может быть частью экзона, который в свою очередь используется в двадцати различных белках. В то же время эта же буква может быть частью кода сплайсинга, который говорит клетке, когда нужно производить каждый из этих белков. Эта буква также может быть частью гистонового кода, который клетка использует, чтобы узнать, в каких местах обвить ДНК вокруг определённых защитных белков, называемых гистонами. Эта буква также может повлиять на трёхмерную структуру ДНК. Она может быть и частью убиквитарного эпигенетического кода (глава 1), и частью трёхбуквенного кодона, который транслируется в определённую аминокислоту. Поскольку геном является четверичным кодом, а также благодаря избыточности кодона, Создатель мог выбирать из альтернативных кодонов, сталкиваясь с необходимостью выполнения одновременно нескольких требований. Избыточность позволила Ему накладывать друг на друга несколько геномных команд без ущерба для требуемой структуры белков.
Существование различных перекрывающихся кодов ДНК и РНК бросает вызов натуралистическому объяснению и не позволяет естественному отбору быть действующей силой долгосрочных эволюционных изменений. Отбор врезается в «стену непреодолимой трудности», когда сталкивается с мутациями, которые затрагивают более чем один признак одновременно. Полифункциональность (также называемая плейотропией) означает, что определённая мутация может повлиять на совершенно не связанные признаки (скажем, цветовое зрение, способность переносить чеснок и митохондриальную эффективность, хотя это было бы крайним примером). Это очередная ахиллесова пята эволюционной генетики. Как мог простой процесс проб и ошибок, который всегда ищет самый простой ответ на вызов окружающей среды, создать перемежающуюся и многоуровневую систему регуляции? Эта система действительно является одним из чудес Вселенной. Без этого уровня многозадачности геном должен был бы быть намного бо́льшим, а возможно, основанные на ДНК многоклеточные организмы вообще не могли бы существовать без него.
Подтверждение книги Бытия в наших генах
Современная генетика открыла нам потрясающе сложный мир, но, кроме этого, генетика также даёт нам возможность проверить различные исторические теории. Существует множество различных древних мифов о сотворении, и разве не удивительно, что только один из них великолепно вписывается в ту картину, которую нам нарисовала генетика? Разве не удивительно узнать об изобильных свидетельствах Сотворения, Потопа и Вавилонской башни в наших генах?
Поскольку Бытие позиционирует себя как книга историческая, и поскольку оно претендует на всеобъемлющую историю человечества, то оно позволяет сделать конкретные и определённые прогнозы относительно генетики человека. Эти прогнозы связаны с сотворением пары первых людей (Адама и Евы), с популяционным бутылочным горлышком, произошедшим около 1600 лет спустя во времена Ноя, когда популяция людей сократилась до восьми душ, и с однократным расселением человеческой расы из одной исходной точки на Ближнем Востоке несколькими веками позже. Мы должны с осторожностью подходить к этому вопросу, поскольку наука о человеке имеет довольно плохой послужной список неправильных выводов, сделанных на протяжении последних столетий. Есть много того, в чём мы можем и сегодня заблуждаться.
Однако, при тщательном и критическом анализе имеющихся данных можно отчётливо увидеть отражение библейской концепции в наших генах. Хотя такие люди, как Френсис Коллинз, руководитель проекта «Геном человека», позиционирующий себя евангельским христианином, и утверждают, что существованию Адама и Евы нет подтверждений,135 на самом же деле похоже на то, что большинство из этих людей слабо либо вообще не представляют, в чём же в действительности заключаются прогнозы библейской модели.
Библия утверждает, что всё человечество произошло от единственной пары. Это очень важное утверждение с точки зрения генетики, поскольку оно существенно ограничивает степень генетического разнообразия людей, наблюдаемую сегодня. Но давайте не будем, подобно эволюционистам, заклиниваться только на моделях, предполагающих начало в необозримом прошлом. Любые потомки Адама и Евы должны быть связаны с ними через естественный процесс половой рекомбинации и воспроизведения, но это не обязательно верно для первоначальной пары. Бог мог вложить в яички Адама различные клеточные линии. Точно также Он мог вложить в яичники Евы различные человеческие геномы, совершенно разные. Первоначальная популяция могла быть удивительно разнообразной. И наоборот, мог быть всего один первоначальный геном, и Ева – практически клон Адама (без Y-хромосомы), возможно даже, гаплоидный клон. Поэтому библейские модели генетической истории человека могут иметь различные вариации. Какую из них выбрать?
По теологическим причинам я предпочитаю версию, что Ева была клоном Адама. Таким образом, она попала под проклятие, наложенное Богом на Адама и его потомков, а также она имеет родство со Христом, являясь кровным родственником Спасителя.136 Также, поскольку она была создана из ребра Адама, можно предположить, что для её создания были использованы клетки, мышцы, нервы, кровеносные сосуды и ДНК (!) Адама. Это не утверждается в Библии, и я открыт к альтернативным объяснениям, однако, склоняюсь к этому варианту.
Возможно, это вас удивит, особенно если до этого вы читали или слышали Френсиса Коллинза, но величина генетического разнообразия, которую мы наблюдаем сегодня, может прекрасно поместиться в Адама и Еву. На самом деле, большая его часть может поместиться в одного Адама! Есть около 10000000 мест, в которых мы находим часто встречающиеся вариации в геноме человека. Средний человек несёт где-то около двух или трёх миллионов из них. То есть существует около трёх миллионов мест, где сестринские хромосомы (хромосомы идут парами; у вас есть, например, две копии хромосомы 1) имеют различные коды в соответствующих местах. Не будет натяжкой считать, что Адам нёс почти все из этих типичных вариаций в своём геноме. Почему люди несут только подмножество из них? Вероятно, это из-за рекурсии в своих родословных. То есть они унаследовали идентичные копии различных участков генома от одного и того же далёкого предка по различным линиям. Добавьте к этому процессы роста, сокращения, разделения и смешения населения в течение библейского времени,137 и нам уже не нужны миллионы людей, живших в далёком прошлом, чтобы объяснить текущее генетическое разнообразие людей.
Кроме того, наиболее типичные геномные вариации существуют в двух версиях. Если же версий оказалось больше двух, то это можно объяснить ранней мутацией (например, наиболее распространённый вариант группы крови O – явно мутация [повреждённая копия] гена, который кодирует аллель группы крови А,138 и эта группа распространена по всему миру) либо мутацией, которая произошла после Вавилонского рассеяния (например, ген серповидно-клеточной анемии, распространённый в некоторых частях Африки, или аллель голубоглазости, распространённый в Северной Европе, а также множество вариаций, определённых генов иммунной системы, которые предназначены быстро меняться). Итак, поскольку большинство вариаций могут поместиться в одного человека, возможно, нет необходимости прибегать к экзотическим моделям ранней библейской генетики.
В рамках «африканской» модели эволюционисты говорят, что человечество прошло через бутылочное горлышко практически полного исчезновения, прежде чем наша популяция разрослась и затем покинула Африку. Почему бутылочное горлышко является частью их модели? Потому что они пытаются объяснить отсутствие разнообразия среди людей, рассеявшихся по всей земле.139 Это разнообразие намного меньше, чем они предполагали изначально, основываясь на версии об огромной популяции, жившей в Африке в течение около миллиона лет. Бутылочное горлышко – это ad hoc дополнение к эволюционной теории, но невысокое разнообразие – идея, изначально присущая креационистской модели.
Кстати, что интересно, с животными всё обстоит иначе. Люди исключительно однообразны, если сравнивать практически со всеми видами животных. Шимпанзе в пять – шесть раз более разнообразны. Обычная комнатная мышь имеет огромный диапазон вариаций в своей популяции, в том числе несколько хромосомных рекомбинаций. Животный мир изобилует подобными примерами. Некоторые из них появились из-за деградации геномов животных со временем. Другие – из-за различных стартовых позиций у животных в сравнении с людьми. Библия не говорит, что Бог создал по две особи каждого рода животных. Фактически, можно предположить, что Он создал уже функционирующую всемирную экосистему, обладающую высоким разнообразием внутри каждого рода (о чём также свидетельствует запись окаменелостей). Это правда, что на Ноевом Ковчеге спаслись лишь по две особи большинства родов животных, но, в отличие от людей, они были выбраны из потенциально намного более обширного генофонда. Поскольку бо́льшая часть генетических данных, имеющихся на сей день, относится к геному человека, нам ещё далеко до возможности построения модели генетической истории большинства видов животных, хотя это интересное направление для будущих исследований.
Помимо множества распространённых вариаций в человеческом геноме, есть также бессчётное количество редких вариаций. Они, как правило, происходят только в изолированных популяциях и указывают на мутации, которые произошли со времён Адама и, конечно, после Вавилонского рассеяния. Они включают в себя, помимо голубых глаз у европейского населения, серповидно-клеточной анемии у африканского населения, также множество сверхредких вариаций, которые встречаются только в отдельном племени, семье или у отдельного человека. Огромное количество генетической изменчивости у нас общее. Это признак того, что мы произошли от небольшой популяции в недавнем прошлом. Те вариации, которые не являются общими на уровне популяции, свидетельствуют о том, что наши геномы быстро деградируют, с высокими темпами мутаций, происходящих во всех этнических группах во всём мире. Подробнее об этом ниже.
Если учесть специфические для женщин и мужчин участки ДНК, мы можем увидеть ещё больше подтверждений Адаму и Еве. Теоретически и практически установлено, что митохондрии (небольшие субклеточные энергетические фабрики, которые преобразовывают сахар в энергию) передаются только по женской линии. Поскольку митохондрии также имеют свой собственный небольшой геном, состоящий примерно из 16569 букв, и поскольку этот маленький геном также подвержен мутациям со временем, мы можем использовать его для построения всемирного генеалогического древа по женской линии. Это привело к появлению в эволюционной литературе ещё в 1987 году гипотезы об африканской митохондриальной Еве.140 Если другие женщины и жили тогда, миллионы лет назад, то только одна сумела передать свой геном всем людям, живущим сегодня.
Считается, что митохондриальная Ева жила сотни тысяч лет тому назад, но только если предполагать определённую медленную скорость мутаций и общее с шимпанзе происхождение. Использование реальной частоты мутаций даёт дату появления Евы около 6000 лет назад.141 Недавнее исследование показало, что мутации в митохондриальной области управления (на них приходится примерно 2/3 всех митохондриальных мутаций) происходят с частотой один раз в поколение.142 Поскольку митохондриальная последовательность Евы была реконструирована и опубликована (в эволюционной литературе)143 и так как большинство митохондриальных линий расходятся менее чем на 30 мутаций от консенсусной последовательности «Евы», а наиболее расходящиеся отличаются всего на 100 мутаций, то разнообразие митохондриальной ДНК в современной человеческой популяции может легко поместиться во временные рамки 6000 лет (200 поколений).
Гипотеза африканской Евы и эволюционные предположения, стоящие за ней, были рассмотрены ранее,144 поэтому мы не будем приводить здесь детали. Известна последовательность более чем 99% митохондриального генома первоначального человека женского пола. Нет никаких данных, указывающих на то, что был кто-либо ещё. Почему люди верят, что Ева была лишь одной из многих женщин, живших в большой популяции давным-давно? Потому что это – часть модели эволюционной истории. Она не основана на реальности, но даёт им удобное оправдание для дискредитации прогноза, основанного на Библии, что генеалогия всех людей в мире по женской линии должна сходиться к единственной женщине.
Подобно митохондриальному геному, мужская Y-хромосома даёт нам возможность построить генеалогическое дерево всех мужчин на Земле. Y-хромосомный Адам якобы существовал также очень давно, но в разное с митохондриальной Евой время. Однако, как и в предыдущем случае, эти выводы основаны на моделях, которые делают массу предположений о человеческой истории, численности населения и частоте мутаций.
После публикации пересмотренной Y-хромосомы шимпанзе145 и открытия, что она только на 70% идентична человеческой Y-хромосоме (даже эта цифра является завышенной, так как половина Y-хромосомы шимпанзе отсутствует), эволюционисты вынуждены были заключить, что эта хромосома мутировала чрезвычайно быстро на протяжении человеческой истории. Однако, современные Y-хромосомы мужчин по всему миру очень похожи друг на друга. Единственный способ сохранить сходство при высоком уровне мутаций – это иметь совсем недавнего общего предка. Viva Адам!
Ещё один способ, которым мы можем воспользоваться, чтобы проверить правдивость библейской истории об Адаме и Еве, – это данные сцепленного наследования. При половом размножении клетки обоих родителей проходят через процесс мейоза, при котором хромосомы, унаследованные от их родителей, рекомбинируются. То есть, когда родители передают свои гены, на самом деле они передают рекомбинированные версии хромосом бабушек и дедушек. Эта рекомбинация, называемая кроссинговером, приводит к тому, что ДНК наследуется крупными блоками. Есть участки генома, которые не рекомбинировались за всю историю человечества (что указывает на молодость генома). Когда два аллеля наследуются вместе (потому что находятся близко друг к другу на одной нити ДНК), их называют сцепленными. Сцепленное наследование было детально изучено, и благодаря ему мы узнали некоторые очень интересные вещи. Во-первых, большинство блоков имеют от двух до четырёх типичных вариантов среди всех человеческих популяций.146 Другими словами, было лишь несколько оригинальных хромосом, и куски этих хромосом остаются нетронутыми. Геном содержит лишь около десяти тысяч блоков, что легко объясняется, если возраст человеческой популяции составляет всего около 200 поколений и за поколение происходят 1–2 кроссинговера на длинное плечо хромосомы.
Помимо предположения о молекулярных часах, стоящего за большей частью эволюционных исследований, большинство также предполагает, что рекомбинация происходила одинаково в ходе всей истории и по всей земле. Но это не обязательно так, ведь, как мы знаем, кроссинговер зависит от генетических факторов (в частности, от гена PRDM9), и существуют различия в этих факторах, которые влияют на частоту кроссинговера у разных людей.147 Это проблема для многих прежних эволюционных исследований, в том числе для тех, на которых основывались подтверждения наших африканских корней.
Из Африки или из Вавилона?
Выше мы обсудили «африканскую» теорию в контексте митохондриальной Евы.тВ этом разделе и давайте просто перечислим соответствия между «африканской» теорией и книгой Бытия. Согласно наиболее популярной эволюционной истории, мы произошли из небольшой популяции, которая распалась на более мелкие ветви в ходе однократного события рассеивания, после которого человечество распространилось по всему миру. Это рассеивание произошло с тремя главными женскими и одной главной мужской линиями в недалёком прошлом. Ах, да, оно прошло через Ближний Восток, прежде чем добралось до Европы, Азии, Австралии, Океании, Северной и Южной Америки. Каждый из этих пунктов предсказывается историей Бытия о Потопе (Быт.6–8), Вавилонской башне (Быт.11) и Таблице Наций (Быт.9–11). Различия во временных рамках (соответственно 6000, 4500, и ~ 4000 лет назад по сравнению с десятками тысяч лет тому назад) и исходной точке (на Ближнем Востоке против северо-восточной Африки около Красного моря148).

Карта миграции человечества, построенная на данных митохондриальной ДНК исходя из «африканской» гипотезы, имеет много параллелей с Книгой Бытия.
Однако выводы «африканской» модели делаются исходя из эволюционных предположений.149 Вкратце, они предполагают, что действовали молекулярные часы, в результате чего мутации накапливались во всех популяциях с одинаковой скоростью на протяжении всего времени. Как мы уже увидели, это не так. Они предполагают, что все группы населения имеют примерно одинаковые демографические показатели (уровни рождаемости и смертности, средний возраст вступления в брак, среднее число детей и т.д.). Они также предполагают, что нет никаких различий в механизмах ремонта ДНК между популяциями, поскольку это нарушило бы ход молекулярных часов. Таким образом, когда в Африке обнаруживают большее разнообразие, автоматически делается вывод, что это более старая популяция и источник остального человечества. Но что, если некоторые африканские племена имеют другую генетическую историю, чем остальные? Есть много африканцев, которые вписываются в средний типичный по миру пул митохондриальной последовательности. Есть другие, которые отличаются гораздо сильнее. Означает ли это, что у них более старые последовательности? Возможно, эти люди по каким-либо другим причинам приобрели больше мутаций в митохондриальной гаплогруппе? Интересно, что по утверждению недавнего исследования, древние африканские племена оставались мелкими и изолированными друг от друга в течение тысяч лет.150 Это может объяснить наблюдаемое быстрое накопление мутаций и генетический дрейф.
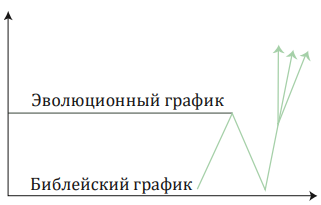
Стилизованное представление «африканского» сценария по сравнению со стилизованным библейским. Эволюционисты полагают, что человек прямоходящий (Homo erectus) существовал в Африке около миллиона лет. Затем по какой-то неизвестной причине вид практически вымер. Современному человеку каким-то образом удалось эволюционировать в этом генетически катастрофическом процессе (катастрофическом, потому что близкородственное скрещивание – это очень и очень плохо). Затем некоторые из новых генетических линий эволюционировали (посредством мутаций) и покинули Африку, распространившись и заселив весь земной шар. Эти гаплогруппы оказались все тесно связанными, и никто не знает, почему другие остались в Африке. Согласно библейской модели, популяция началась с Адама и Евы, увеличилась до неизвестного числа, через 1 600 лет уменьшилась до восьми человек и вновь начала расти. Затем вся популяция распространяется по всему миру.
Есть некоторые дополнительные параллели между «африканской» моделью и Книгой Бытия. Как правило, считается, что люди существовали в Африке несколько миллионов лет как человек прямоходящий (Homo erectus). Затем, без объяснения причин, эта досовременная популяция людей пережила катастрофу и практически вымерла. Десяти тысячам или менее удалось выжить, и они быстро эволюционировали в современного человека. Население восстановилось, диверсифицировалось, а затем некоторым генетическим линиям удалось покинуть Африку и покорить мир. Библейская история начинается с двух людей, Адама и Евы. Численность популяции растёт до некоторого неизвестного количества и затем при Потопе, произошедшем через 1600 лет после Сотворения, падает до восьми душ, среди которых три способных к воспроизводству пары. Популяция людей затем вновь увеличивается, но они снова бросают вызов Богу, как это было до Потопа, Он вмешивается и смешивает их языки в месте под названием Вавилон, заставляя их разбиться на кланы и разойтись по разным сторонам света. Таким образом, они рассредоточиваются и покоряют мир после происшествия с Вавилонской башней, случившегося спустя несколько поколений после Потопа. Существует хорошее общее соответствие между «африканским» сценарием и Книгой Бытия, после того как эволюционная история была модифицирована, чтобы согласовать её с данными. В частности, «африканской» модели приходится иметь дело с отсутствием разнообразия среди людей во всём мире (а значит, с сокращением популяции) и единоразовым рассеиванием человечества, что проявляется в наших генах.
Удивительный неандерталец
Как нам быть с утверждениями о наших нечеловеческих пещерных предках? Новые открытия в археологии и генетике заставили эволюционные взгляды на неандертальцев радикально измениться буквально за одно последнее десятилетие. Сегодня считается, что неандертальцы рисовали в пещерах, создавали музыкальные инструменты, использовали огонь, церемониально хоронили своих умерших головой в сторону рассвета и искали на местности редкие минералы для того, чтобы растереть их и использовать для макияжа (косметики).151 Детали этого списка всё ещё обсуждаются между различными эволюционистами, но даже одна единственная из них, не говоря уже о нескольких одновременно, была бы равносильна эволюционной ереси всего несколько лет назад.
Благодаря стремительному технологическому прогрессу, сейчас у нас есть возможность получить ДНК из некоторых лучше всего сохранившихся костей неандертальцев. Область генетики древней ДНК сталкивается с определёнными трудностями, так как ДНК – это хрупкая молекула, которая быстро распадается после смерти организма. Кроме того, некоторые из повреждений подобны тем, которые происходят со временем в живых организмах. Поэтому иногда трудно отличить посмертный распад ДНК от мутаций, которые произошли в родословной этого человека. Ещё одной проблемой является загрязнение. Поскольку ДНК древнего образца неизбежно является деградировавшей, то любое загрязнение со стороны современной ДНК будет заглушать данные по древней ДНК. Исследователи хорошо знают об этих проблемах и приложили много усилий, чтобы преодолеть их, в частности, они работают с любой новой находкой как будто на месте совершения преступления, в том числе используя метод «чистой комнаты», предназначенный для уменьшения загрязнения людьми, работающими с костями.152

Буквами показаны первые 330 аминокислот белка FOXP2 человека, шимпанзе, гориллы, орангутанга, резус-макаки и мыши. Последовательности аминокислот содержат два полиглютаминовых участка (обозначены красным) и две специфические мутации (нижний ряд), которые отличают последовательность человека от остальных представленных млекопитающих (N на позиции 304 и S на позиции 326). Концевые 386 аминокислот FOXP2 идентичны у всех видов и здесь не показаны. Последовательности приведены согласно Enard и др. Enard W. и др., Molecular evolution of FOXP2, a gene involved in speech and language, 418:869–872, 2002 г.
Что же мы выяснили? Удивитесь ли вы, узнав, что генетика привела к ещё одному неожиданному для эволюционистов обстоятельству? После тщательного рассмотрения вопросов, перечисленных выше, современные исследования по генетике неандертальцев привели к поразительным выводам. На начальном этапе было обнаружено, что неандертальцы имели тот же ген (FOXP2), который даёт современным людям возможность говорить.153 Затем было обнаружено, что некоторые неандертальцы имели подобные версии генов пигмента кожи, определяющие светлую кожу и рыжие волосы, а также зелёные глаза и веснушки, которые встречаются у людей европейского происхождения.154 Всё указывало на то, что они были людьми в гораздо большей степени, чем считалось ранее. Но потом на основе митохондриальной ДНК, извлечённой из образцов ткани неандертальцев, был сделан вывод, что они, вероятно, не скрещивались с предками современного человека, потому что митохондриальные последовательности неандертальцев у людей, живущих сегодня, найдены не были.155
Эти выводы, однако, оказались скоротечными, потому что вскоре был опубликован полноразмерный геном неандертальца (если быть точными, 60% от него).156 Если последовательности точны, то неандерталец является не тем, кем его ожидали увидеть. Сейчас существуют подтверждения тому, что неандертальцы скрещивались с прямыми предками современных людей, то есть, согласно биологической концепции вида, мы – один и тот же вид. Похоже, что люди, живущие за пределами Африки, несут 3–4% ДНК неандертальцев. Интересно, что останки неандертальцев не были найдены в Африке, но следы их генома всё ещё сохраняются в тех местах, где они когда-то жили (и не только). Новое эволюционное объяснение – это модифицированный «африканский» сценарий, согласно которому происходило ограниченное скрещивание между современными людьми и неандертальцами, когда первые покинули Африку и вытеснили неандертальцев на своём пути к мировому господству. Но это радикально меняет статус-кво, существовавший ещё совсем недавно. Помните, какими уверенными эволюционисты были, когда талдычили библейским креационистам про гипотезу африканского происхождения? Многое из того, в чём они были уверены, в настоящее время не подкрепляется их собственными научными данными.157
Существует альтернативный вариант, который также соответствует данным. Можно предположить, что вместо двух больших групп населения, которые ограниченно смешивались, популяция неандертальцев была намного меньше, чем основная волна миграции, и возможно, даже было полное перемешивание, что привело к тому же результату. Если доля неандертальской ДНК в современных неафриканских популяциях составляет 3–4%, то, возможно, их численность как раз и была в пределах 3–4% от размера популяции современных людей или где-то в этих пределах при условии другой степени смешения. Если неандертальцы были людьми, особенно если это была послепотопная популяция, смешивание было естественным, ведь люди, в конце концов, делают то, что делают. Более поздние данные показали, что неандертальцы по всей территории своего обитания были более похожи друг на друга, чем индивидуумы в любой современной группе людей.158 От Испании и до центральной России неандертальцы больше похожи на одно большое семейство – семейство людей, которые жили в Европе и Азии после Потопа и были окончательно вытеснены более поздней волной миграции.
Человек и шимпанзе
В течение нескольких десятилетий мы слышали: «Люди и шимпанзе на 99% идентичны». Это не правда.159 Эта цифра основана на некоторых ранних экспериментальных данных, полученных путём сравнения участков известных генов между собой, так что по крайней мере некоторая часть наших ДНК очень похожа. Но лишь менее чем 2% наших геномов кодируют белок, а многие гены двух видов невозможно сравнить. Люди имеют несколько сотен кодирующих белки генов (все они плотно интегрированы в сплайсосоме), которые отсутствуют у шимпанзе. У человека найдены целые семейства генов, которых нет у шимпанзе.160 Это вставляет палки в колёса эволюционных моделей, поскольку с момента, когда мы предположительно были одним видом, прошло всего около нескольких сотен тысяч поколений.161 Каким образом эти совершенно новые гены могли возникнуть и затем интегрироваться в наши сложнейшие геномы за такой короткий период времени? Время – не решающий фактор для эволюции. Эволюция измеряется в поколениях, а их прошло не так много от предполагаемого общего предка.
Фактически, есть около 35000000 однобуквенных различий,162 которые должны были возникнуть (половина в каждом из видов), распространиться в соответствующей популяции и стать фиксированными (изначальная буква на этой позиции должна полностью исчезнуть из популяции) за это небольшое количество поколений. Аналогично, десятки тысяч хромосомных рекомбинаций должны были произойти, распространиться и зафиксироваться, так же как и десятки миллионов пар оснований вставок и удалений. Скорость, с которой могут происходить фиксации, – медленная, и большинство новых вариантов должны были исчезнуть (поскольку они по определению редки). Скорость мутаций и отбора, необходимая для такого объёма изменений за всего лишь 6 миллионов лет, ошеломляет, но они должны были произойти, чтобы эволюция была правдой.
Отсутствие сходства между Y-хромосомами шимпанзе и человека было описано выше. С учётом всего лишь 70% сходства с одной половиной Y-хромосомы шимпанзе, которая существует, частота мутаций должна быть чрезвычайно высокой. Почему же у мужчин настолько одинаковые хромосомы? Возможно, потому, что хромосоме всего 6000 лет!
Люди и шимпанзе ожидаемо несколько подобны. Шимпанзе выглядят похоже на нас, они ведут себя во многом, как мы, они едят те же продукты, что и мы, и имеют схожие с нами экологические потребности (за исключением того, что мы были достаточно умны, чтобы изобрести тёплые дома и тёплые вещи). С какой стати ожидать, что они не будут похожи на нас на генном уровне? Представьте себе, что бы говорили эволюционисты, если бы никакого сходства не было? Они бы утверждали, что нет доказательств разумного Творца, так как следовало бы ожидать, что Творец использует общий шаблон дизайна для подобных организмов. Они в выигрыше в любом случае!
| Источник | Общее количество проанализированных геномных оснований | Совмещённых оснований | Указанное совпадение ДНК | Фактическое совпадение ДНК163 |
| Britten, 2002 г. | 846016 | 779132 | 95,2% | ~ 87% |
| Ebersbergerи др., 2002 г. | 3000286 | 1944162 | 98,8% | < 65% |
| Liu и др., 2003 г. | 10600000(суммарно почеловеку, шимпанзе, бабуину и мартышке) | 4968069(человек – шимпанзе) | 98,9%не включаявставки и удаления | ? |
| Wildmanи др., 2003 г. | ~90000(экзоны 97 генов) | ? | 98,4–99,4% | ? |
| Chimp. Chrom. 22 Consort. | 32799845 | ? | 98,5%не включая вставки и удаления | 80–85%включая вставки и удаления |
| Nielson и др., 2005 г. | ?164 | ? | 99,4%избранные участки генов | ? |
| Chimp. Seq. Consort. 2005 г. | Целый геном(5-кратное покрытие) | 2,4 Гб | 95,8% | 81%165 |
Обзор работ по сравнению геномов человека и шимпанзе. Где это возможно, отброшенные при сравнении данные использованы для расчёта фактического процента сходства ДНК (согласно Tomkins и Bergman, 2012 г.)166.
Насколько мы похожи? Как сказал известный генетик Сванте Паабо:
Я не думаю, что есть какой-то способ посчитать конкретную цифру … в конце концов, то, как мы видим наши отличия, – политический, социальный и культурный вопрос.167
Темпы мутаций слишком высокие
В главе 1 рассматривались влияние высоких темпов мутации и тот факт, что естественный отбор не может «видеть» большинство мутаций. Вывод заключался в том, что большинство мутаций, несмотря на то, что они вредны, проскальзывают сквозь сито отбора. Это означает, что вредные мутации накапливаются в наших видах, а это является противоположностью тому, чего требует длительная эволюция. Примените эту мысль к нашему современному пониманию сложности генома, и вы начнёте видеть масштабы проблемы. Геномы эукариотов (все организмы, кроме бактерий) слишком сложны, и мишень для мутаций слишком велика, чтобы эволюция могла работать. Средняя буква копируется на шесть различных транскриптов РНК и используется в нескольких перекрывающихся кодах (гистонный код, белковый код, сплайсинг и т.д.). Мутации происходят с ошеломляющей для эволюционистов скоростью, устанавливая предел долголетия всей человеческой расы. Однако ясно одно – Иисус обещал вернуться, и Он пообещал, что люди всё ещё будут здесь, когда Он вернётся. Таким образом, мы не вымрем до Второго пришествия. Когда оно будет? На основании прогнозируемых вредных долгосрочных последствий, причиняемых геному мутациями, конечно, не миллионы лет спустя.
Вызов
Геном – это многомерная операционная система со встроенными кодами для исправления ошибок и самомодификации. В нём присутствуют различные перекрывающиеся ДНК-коды, РНК-коды и структурные коды. Существуют ДНК-гены и РНК-гены.
Геном был специально спроектирован с высокой степенью избыточности. Несмотря на избыточность, он демонстрирует поразительную степень компактности, так как около 20000 «генов» комбинаторно создают намного более 100000 различных белков. Также геном медленно вырождается с течением времени, однако он до сих пор остался жизнеспособным благодаря прекрасно спроектированным помехоустойчивым кодам и встроенной, разумно спроектированной избыточности.
Известная цитата Дарвина звучит так:
Если бы можно было показать, что существует какой-либо сложный орган, который не мог никак сформироваться путём многочисленных последовательных небольших изменений, моя теория, безусловно, разрушилась бы.168
Я знаю, что эта цитата в течение многих лет использовалась некорректно (обеими сторонами), но всё-таки я утверждаю, что геном человека как раз отвечает такому критерию. Я не верю, что он мог произойти путём естественных процессов. И я бросаю вызов эволюционистам: представьте работоспособный сценарий истории генома, включающий источник информационных изменений, объяснение количества необходимых мута- ций, описание необходимых факторов отбора, и всё это в соответствующих временных рамках. Фактически, я ставлю перед ними сложную задачу описать, как геном, любой геном, мог возникнуть с нуля в мире, в котором отсутствовала содержательная информация.
Куда это ведёт?
Сейчас должно быть очевидно, что генетика – не союзник Дарвина. Его неосведомлённость о сложности жизни, о способах, с помощью которых виды воспроизводят себя, и о хрупкости сложных систем позволили ему беспрепятственно теоретизировать. Дарвинизм нужно оценить в свете современных знаний. Современная генетика довольно хорошо поддерживает библейскую историю. Гены современного человека содержат множество свидетельств существования двух первоначальных людей (Адама и Евы), бутылочного горлышка несколько тысяч лет спустя (во время Ноева Потопа) и разделения популяции несколькими поколениями позже (в Вавилоне) с последующим единоразовым рассеянием человечества по земному шару. Кроме того, скорость мутаций, распределение мутаций, и хрупкость ультрасложной компьютерной операционной системы под названием «человеческий геном» – всё это свидетельствуют о молодости этой системы. Интересно, мог ли Дарвин прийти к таким же выводам, если бы он жил сейчас (и если бы он открыто и честно проанализировал соответствующие данные). Однако, не только генетика является противником эволюции, само происхождение жизни также не является союзником эволюционной теории. В следующей главе мы рассмотрим этот вопрос, поскольку все эволюционные сценарии происхождения жизни нарушают известные законы химии, физики и вероятности.
* * *
Darwin, C.R., The variation of animals and plants under domestication, Лондон, John Murray, 1868 г.; darwin-online.org.uk.
Carter, R., Darwin’s Lamarckism vindicated?, март 2011 г.; creation.com/epigenetics-and-darwin.
Druery, C.T. и Bateson, W., Experiments in plant hybridization, Journal of the Royal Horticultural Society 26:1–32, 1901 г.
Отец послал его учиться в Кембридж после того, как он бросил (или был отчислен) обучение в медицинской школе в Эдинбурге. Окончив Кембридж, он отправился в своё знаменитое пятилетнее кругосветное путешествие на корабле Бигль и никогда уже не возвращался к мысли стать священником.
Мендель показал, что при скрещивании двух организмов, оба из которых несут доминантные и рецессивные версии гена (например, доминантные фиолетовые лепестки и рецессивные белые лепестки), получается соотношение 3:1 доминантного типа (фиолетовый) и рецессивного (белый). Это происходит потому, что белые цветы будут только у растений, которые получат две копии рецессивного гена. Таким образом, 3/4 растений будут иметь фиолетовые цветы и 1/4 – белые, то есть соотношение 3:1 фиолетового и белого.
Howard, J.C., Why didn’t Darwin discover Mendel’s laws? Journal of Biology 8:15, 2009 г
1. В том числе: The Variation of Animals and Plants under Domestication, 1868 г.; The Effects of Cross and Self-Fertilisation in the Vegetable Kingdom, 1876 г.; и The Different Forms of Flowers on Plants of the Same Species, 1877 г.
Howard, J.C., ссылка 6.
Lodish, H. и др., Molecular Biology of the Cell, 5-е изд., W.H. Freeman and Company, Нью-Йорк, 2004 г.
Batten, D., Book review: The Biotic Message: Evolution versus Message Theory, J. Creation 11(3):292–298; creation.com/biotic.
Gitt, W., Scientific laws of information and their implications – part 1, J. Creation 23(2):96–102; сreation.com/laws-of-information-1. См. также другие части этой серии статей.
Lynch, M., Rate, molecular spectrum, and consequences of human mutation, Proc. Nat. Acad. Sci. USA 107(3):961–968, 2010 г. Похожие цифры были получены во многих других исследованиях, в том числе Neel, J.V. и др., The rate with which spontaneous mutation alters the electrophoretic mobility of polypeptides, Proc. Nat. Acad. Sci. USA 83:389–393, 1986 г.; Nachman, M.W. и Crowell, S.L., Estimate of the mutation rate per nucleotide in humans, Genetics 156:297–304, 2000 г.; Kondrashov, S., Direct estimates of human per nucleotide mutation rates at 20 loci causing Mendelian disease, Human Mutation 21:12–27, 2002 г.
Eyre-Walker, A. и Keightley, P.D., High genomic deleterious mutation rates in hominids, Nature 397:344– 347, 1999 г.
Lynch, M., Rate, molecular spectrum, and consequences of human mutation.
Crow, J.F., The origins, patterns, and implications of human spontaneous mutation, Nature Reviews: Genetics 1:40–47.
Conrad, D.F. и др., Variation in genome-wide mutation rates within and between human families, Nature Genetics 43:712–714, 2011 г.
Kondrashov, A.S., Contamination of the genome by very slightly deleterious mutations: why have we not died 100 times over, J. Theor. Biol. 175:583–594.
ReMine, W.J., Cost theory and the cost of substitution–a clarification, J. Creation 19(1):113–125, 2005 г.; creation.com/cost.
Haldane, J.B.S., The cost of natural selection, Journal of Genetics 55:511–524, 1957 г.
Batten, D., Haldane’s dilemma has not been solved, J. Creation 19(1):20–21, 2005; creation.com/Haldane. См. также saintpaulscience.com/Haldane.htm.
Kimura, M., Evolution rate at the molecular level, Nature 217:624–626, 1968 г.
ReMine, W.J., ссылка 18.
Ohno, S., So much «junk» DNA in our genome, Evolution of genetic systems, Brookhaven Symposia in Biology, no. 23 (под ред. Smith, H.H.), стр. 366–370, 1972 г.
Carter, R.W., The slow, painful death of junk DNA, J. Creation 23(3):12–13, 2009 г.; creation.com/ junk-russian.
Birney, E. и др., Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project, Nature 447:799–816, 2007 г.
J.S. Mattick, цит. по Gibbs, W.W., The Unseen genome: gems amid the junk, Scientific American, стр. 47– 53, ноябрь 2003 г.
Yan, K.-K. и др., Comparing genomes to computer operating systems in terms of the topology and evolution of their regulatory control networks, Proc. Natl. Acad. Sci. USA 107(20):9186–9191, 2010 г.
Yan, K.-K. и др., Comparing genomes to computer operating systems in terms of the topology and evolution of their regulatory control networks, Proc. Natl. Acad. Sci. USA 107(20):9186–9191, 2010 г.
Stein, L.D., Human genome: end of the beginning, Nature 431:915–916, 2004 г.
Birney, E. и др., ссылка 25; см. также Williams, A., Astonishing DNA complexity update, июль 2007 г., creation.com/dnaupdate.
Barash, Y. и др., Deciphering the splicing code, Nature 465:53–59, 2010 г.
См. Anon., Human genes sing different tunes in different tissues, PhysOrg.com, 2 ноября 2008 г.
Carter, R.W., Splicing and dicing the human genome, июль 2010 г.; creation.com/splicing.
Lynch, M., Rate, molecular spectrum, and consequences of human mutation.
Gerstein, M.B. и др., What is a gene, post-ENCODE? History and updated definition, Genome Research 17:669–681, 2007 г.
Schoenfelder, S. и др., Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells, Nature Genetics 42:53–61, 2009 г.; См. также Scientists’ 3-D view of genes- at-work is paradigm shift in genetics, sciencedaily.com, 16 декабря 2009 г.
Eitan, Y. и Tanay, A., Probabilistic modeling of Hi-C contact maps eliminates systematic biases to characterize global chromosomal architecture, Nature Genetics 43(11):1059–1067, 2011 г.
Is the shape of a genome as important as its content? PhysOrg.com, 29 октября 2010 г.
Bolzer, A. и др., Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes, PLoS Biol 3(5):e157, 2005 г.
См. A new look at how genes unfold to enable their expression, PhysOrg.com, 14 июля 2008 г.
Baillie, J.K. и др., Somatic retrotransposition alters the genetic landscape of the human brain, Nature 479(7374):534–537.
Li, G.-W., Oh, E. и Weissman, J.S., The anti-Shine-Dalgarno sequence drives translational pausing and codon choice in bacteria, Nature 484(7395):538–541, 2012 г.
Например, в двоичной системе было бы необходимо как минимум 5 букв, чтобы закодировать 20 аминокислот, с меньшей избыточностью (25 = 32 варианта).
Carter, R.W., The non-mythical Adam and Eve: refuting errors by Francis Collins and BioLogos, август 2011 г.; creation.com/biologos-adam-russian.
Sarfati, J., The Incarnation: Why did God become Man? Декабрь 2010 г.; creation.com/incarnation-russian.
Carter, R.W., ссылка 43.
Sarfati, J., Blood types and their origin, J. Creation 11(1):31–32, 1997 г.; creation.com/blood-groups.
Carter, R.W., The Neutral Model of evolution and recent African origins, J. Creation 23(1):70–77; creation.com/african-origins.
Cann, R.L., Stoneking, M. и Wilson, A.C., Mitochondrial DNA and human evolution, Nature 325:31–36, 1987 г.
Wieland, C., A shrinking date for eve, J. Creation 12(1):1–3, 1998 г.; creation.com/eve.
Madrigal, L. и др., High mitochondrial mutation rates estimated from deep-rooting Costa Rican pedigrees, American Journal of Physical Anthropology 148:327–333, 2012 г. См. также Carter, R.W., Is ‘mitochondrial Eve’ consistent with the biblical Eve? Январь 2013 г.; creation.com/mteve-biblical-eve.
Carter, R.W., Mitochondrial diversity within modern human populations, Nucl. Acids Res. 35(9):3039–3045, 2007 г.
Carter, R.W., ссылка 43; Carter, R.W., ссылка 47.
Hughes, J.F. и др., Chimpanzee and human Y chromosomes are remarkably divergent in structure and gene content, Nature 463:536–539.
The International HapMap 3 Consortium, Integrating common and rare genetic variation in diverse human populations, Nature 467:52–58, 2010 г.
Parvanov, E.D. и др., PRDM9 controls activation of mammalian recombination hotspots, Science 327:835, 2010 г.; Berg, I.L. и др., PRDM9 variation strongly influences recombination hot-spot activity and meiotic instability in humans, Nature Genetics 42(10):859–864, 2010 г.; См. также, Carter, R.W., Does genetics point to a single primal couple? Апрель 2011 г.; creation.com/genetics-primal-couple.
Tishkoff, S.A. и др., The genetic structure and history of Africans and African Americans, Science 324:1035–1044, 2009 г.
Carter, R.W., ссылка 47.
Behar, D.M. и др., and The Genographic Consortium, The dawn of human matrilineal diversity, Am. J. Human Gen. 82:1130–1140, 2008 г.
Carter, R.W., The Painted Neanderthal, май 2010 г.; creation.com/the-painted-neandertal.
Carter, R.W., Neandertal genome like ours, июнь 2010 г.; creation.com/neandergenes.
Borger, P. и Truman, R., The FOXP2 gene supports Neandertals being fully human, J. Creation 22(2):13– 14; creation.com/foxp2.
Lalueza-Fox, C. и др., A Melanocortin 1 Receptor allele suggests varying pigmentation among Neanderthals, Science 318:1453–1455, 2007 г.
Green, R.E. и др., A complete Neandertal mitochondrial genome sequence determined by highthroughput sequencing, Cell 134:416–426, 2008 г.; Carter, R.W., The Neandertal mitochondrial genome does not support evolution, J. Creation 23(1):40–43, 2009 г.; creation.com/neandertal-mito.
Green, R.E. и др., A draft sequence of the Neandertal genome, Science 328:710–722, 2010 г.
Carter, R.W., Neandertal genome like ours (There may be Neandertals at your next family reunion!), июнь 2010 г.; creation.com/neandergenes.
1. Reich, D. и др., Genetic history of an archaic hominin group from Denisova Cave in Siberia, Nature 468:1053–1060, 2010 г. См. также Wieland, C. и Carter, R.W., Not the Flintstones–it’s the Denisovans, январь 2011 г.; creation.com/denisovan.
Tomkins, J. и Bergman, J., Genomic monkey business–estimates of nearly identical human-chimp DNA similarity re-evaluated using omitted data, J. Creation 26:94–100, 2012 г.; creation.com/chimp.
Demuth J.P. и др., The evolution of mammalian gene families, PLoS ONE 1(1): e85, 2006 г.
6 миллионов лет / ~20 лет на поколение = всего 300000 поколений.
Varki, A. и Altheide, T.K., Comparing the human and chimpanzee genomes: searching for needles in a haystack, Genome Research 15:1746–1758, 2005 г.
На основе размеров отброшенных участков ДНК при совмещении.
Невозможно вычислить фактический процент сходства, потому что отброшенные данные не были предоставлены.
В сравнении с данными The International Human Genome Sequencing Consortium (2004) – ((0,9577 x 2,4 Гб) / 2,85 Гб) x 100
Statham, D., Heads I win, tails you lose: the power of the paradigm, ноябрь 2010 г.; creation.com/fused.
Cohen, J., Relative differences: the myth of 1%, Science 316:1836, 2007 г.
Darwin, C., Origin of Species, 6-е изд., 1872 г.