
Менделизм
Слово „менделизм“ имеет такую же давность, как и двадцатое столетие. Оно введено в науку на рубеже двух столетий – в 1900 году, хотя тот ученый, от имени которого оно произведено – прелатъ Григорий Мендель, – умер еще в 1884 году. Менделизмом называют учение о наследственности предложенное Менделем. Сущность наследственности выражается формулою „подобное производит подобное“ (like begets like), но эта формула дополняется другою: „природа никогда не пользуется дважды одною и тою же формой“ (nature nexer uses the same would twice)24; таким образом подобное производит подобное, но не тождественное. Дети похожи на родителей, но не тождественны с ними. Этот факт порождает рядъ вопросов, решение которых имеет громадное теоретическое и практическое значение. От родителей детям передаются и физические свойства и нравственныя качества. В одних случаях передаются одни, в других – другие; в одних случаях передаются свойства отцовские, в других – материнские. Порою происходит еще нечто более загадочное: передаются свойства деда, бабки и т. д. по восходящей линии и такие свойства, которых у ближайших предков не было. Какие законы управляют этою передачею? Нельзя ли, установивши эти законы, направить наследственность по наиболее целесообразному пути: изменять породы растений и животных в желательном направлении, улучшать род человеческий?
Теорий наследственности было предложено много. Важнейшие из них будут изложены далее. Но хотя эти теории предлагались натуралистами, от них всецело веяло метафизикой. О дарвиновской теории наследственности русский дарвинист Тимирязев сказал, что она ненаучна в основе и безплодна по выводам. Теория монаха Менделя исходит из опыта и утверждается на опыте. Еще в 1865 г. Мендель сделал доклад обществу Брюннских натуралистов о своих восмилетних опытах над растениями. Этот доклад был напечатан в Verhandlungen des naturforschenden Vereines in Brünn (IV, 1865, R. S. 3–57) под заглавием Versuche über Pflanzenhibriden. Въ 1869 г. Мендел еще опубликовал о своих опытах над Hierасіum: ueber einige aus künstlicher Befruchtung gewonnene Hieräcium – Bastarde. Hieräcium – наша полевая ястребинка, сложноцветное растение в ¼–½ фута вышиною, растет по дорогам, имеет желтый дветок, листья и стебель с волосками. Важнейшая работа Менделя über Pflanzenhybriden имеет немного более 40 страниц; кроме названных работ он напечаталъ лишь несколько маленьких заметок и от него остались тепер уже изданные письма к знаменитому мюнхенскому ботанику Негели – Gregor Mendel’s Briefe an Carl Nägeli, herausgegeben von C. Correns. 1905.
До 1900 года работы Менделя оставались неизвестными ученому миру. В этом году их извлек из мрака забвения голландский ботаник Фриз – творец мутационной теории происхождения видов – и одновременно с ним некоторые другие выдающиеся ботаники. Теперь около имени Менделя и его теории ведутся горячие научные споры, производятся опыты, издаются изследования. На русском языке уже в 1909 году явился перевод учебника ботаники Страсбургера, где уделяется внимание менделизму, и перевод экспериментальной зоологии Моргана, в которой менделизму отведено уже большое место. В 1913 году переведены P. К. Пеннета – менделизм (R. C. Punnett – Mende-lism. Cambridge. 1910) и Л. Займемся менделизмом и мы. Но прежде сообщим краткие сведения о Менделе, загробный покой которого слава потревожила лишь десятилетия спустя после его смерти.
Мендель родился в 1822 году в Гейцендорфе, в австрийской Силезии, в семье небогатаго мызника. При крещении он получил имя Иоанна. Благодаря денежной помощи сестры, он окончил курс в гимназии в Топпау. Почувствовав призвание к монашеской жизни, он вступил в августинский орден, принявши имя Григория, поселился в брюннском монастыре Фомы аквинскаго (Брюнн – главный город Моравии); здесь в 1847 году он стал священником.
Благодаря отцу, Мендель с детства был знаком с садоводством и в монастырских садах начал производить опыты над растительными видами. Его начальство послало его для довершения научнаго образования в венский университет, где Мендель пробыл с 1851 по 1853 год. Возвратившись в Брюнн, он стал преподавателем в реальном училище этого города и занимался этим до 1868 года, когда был избран прелатом своего аббатства.
В годы своей преподавательской деятельности он занимался изучением явлений наследственности при скрещивании, главным образом у растений. Его опыты с садовым горохом отняли у него восемь лет. Но он производил опыты не только над растениями, он производил их над пчелами.
Высокий пост занятый Менделем отнял его у науки. Это – история повторявшаяся со многими выдающимися учеными. Это случилось с самим Исааком Ньютоном, который получив пост директора монетнаго двора, в сущности должен был оставить свои научные работы. Мендель с конца шестидесятых годов не дает никаких исследований. Дела по управлению монастырем поглощают его всецело. В начале семидесятых годов ему еще пришлось вести борьбу с австрийским правительством по вопросу о специальном налоге на имущества монашеских орденов. Монастырские сады и огороды были забыты. Вернуться к ним Менделю уже не пришлось. Он умер от брайтовой болезни в 1884 году.
1. Работы Менделя
Мендель поставил своею задачею изучать явления наеледственности при скрещивании. Скрещиванием в животном мире называется спаривание самцов и самок различных пород или разновидностей. Скрещиванием у растений является опыление пестика одной разновидности или породы цветочною пылью другой. ІІри скрещивании вообще происходит соединение индивидуумов различающихся между собою одним или несколькими признаками. Мендель и поставил своею задачею проследить судьбу таких признаков в потомстве.
Половое размножение состоит в том, что женская половая клетка – яйцо или оосфера – оплодотворяется мужской половою клеткою – сперматозоидом или антерозоидом.
ІІоловую клетку – независимо от того мужская она или женская – называют вообще гаметою. Соединение мужскои и женской гаметы, полагающее начало новому индивидуму, называется зиготой. Выяснение свойств гамет и зигот конечно должно служить ключем к пониманию явлений наследственности, но в деле исследования приходится идги обратным путем и от явлений и фактов наследственности делать заключения к свойствам гямегт, и зигот.
Мендель дает следующие методологические правила для экспериментальнаго исследования вопроса.
I. Опыты скрещивания должны быть такими, чтобы они давали возможность наблюдать все формы могущие появиться в потомстве. Нужно выбирать такой материал, скрещивание когораго не влечет за собою никакого уменьшения плодовитости в потомстве. Большее или меньшее безплодие может помешать появлению некоторых наследственных черт в потомстве.
2. Для скрещивания нужно выбирать формы отличающиеяся одна от другой одним или несколькими постоянными и резко различными признаками.
3. Нужно точно определять судьбу родительских признаков у потомства. Такие положения, что потомки ближе к типу отцовскому, к типу материнскому, слишком общи и неопределенны, чтобы из них можно было делать выводы.
4. Необходимо не топько изучать судьбы потомства на многих поколениях, но нужно изучать отдельно судьбу каждой семьи поколения.
5. Законы наследственности можно выводить только из детальных и точных статистических данных. Нужно подвергать наблюдению все потомство каждаго поколения и определять точно число индивидуумов, в когорых проявляется тот или иной наследствевный тип.
6. Наконец, произведя скрещивание раз, никаким образом нельзя допускать, чтобы скрещивание как-либо повторилось в потомстве.
Многое в этих правилах до XX столетия было неизвестно и не принималось в расчет ботаниками и зоологами25.
Для своих опытов Мендель выбрал известный всем съедобный горох (pisum sativum). Разновидностей его очень много. Эти разновидностн отличаются одна от другой ростом (высокий и карликовый горох), формою зерна (круглое, морщинистое, угловатое), цветом семянодолей (желтый, зеленый), цветом оболочки зерна (серая, коричневая, белая) и другими признаками. Менделю согласно его методологическим правилам нужно было для скрещивания выбрать разновидности с резко различающимися признаками. Высокий горох, как и низкий не весь одинаковаго роста. Возможно, что и наиболее низкие экземпляры первого будут равны высшим экземплярам второго. Такие разновидности негодны для опытов. Нужно, чтобы в данной разновидности не было ни одного экземпляра, который не отличался бы изследуемым признаком от всех зкземпляров другой разновидности. Такие разновидности Мендель и выбирал. Простейшая задача поставленная им состояла в том, чтобы проследить судьбу одного признака у потомства происшедшаго от скрещивания. Это – опыты с моногибридами.
Мендель исследовал семь видов скрещивания в Pisum Sativum.
высокий стебель+стебель карликовый
желтые зерна+зеленые зерна
круглые зерна+угловатые зерна
коричневые семеоболочка+безцветная
стручек вздутый+стручек плоский
стручек зеленый+стручек желтый
цветы пазушные+цветы верхушечные.
Для выяснения всего сделаннаго Менделемъ достаточно, сказать об его опытах с высоким и низким горохом.
Он опылял высокорослым горохом низкий н наоборот затем высеял семена происшедшие от скрещивания и получил исключительно высокое потомство без всяких промежуточных форм. Высокорослость Мендель назвалъ признаком доминантным (господствующим), а низкорослость – признаком рецессивным (уступающим). Он снова посеял семена. Выросло второе поколение. ¾ его было высокорослым, ¼ – низкорослым. Первое поколение можно обозначить исключительно буквою Д (доминант), для второго поколения подходит формула: 3D+1R (3 доминанта, 1 – рецессив). Факт, что часть потомства высокорослаго гороха оказалась низкорослою, дает вывод, что свойства индивидуума определяются не только явными его признаками, но еще и теми, которые являются в его потомстве. Потому нужно изолированно изучать потомство второго поколения, тщательно избегая новаго скрещивания. Мендель продолжил опыты. Потомство низкорослаго гороха было исключительно низкорослым во всех последующих поколениях. Судьба высокорослаго гороха была гораздо сложнее.
Низкорослый горох второго поколения, очевидно, владел признаком низкорослости в чистоте; в нем не было ничего, что могло бы противодействовать проявлению этого признака в потомстве. У высокорослого гороха только часть потомства во всех последующих поколениях давало исключительно высокорослые экземпляры, т. е. значит только часть потомства владела признаком высокорослости в чистоте.
Одна треть потомства высокорослого гороха, т. е. значит одна четверт всего потомства, давала исключительно высокорослых индивидуумов. Остальное высокорослое потомство. т. е. две трети его или иначе половина всего потомства, дало частью доминантное, частью – рецессивное поколение, причем взаимоотношение их по численности опять подходило под формулу: 3D + 1R. Так мы получаем: от скрещивания высокорослого гороха с низкорослым получился исключительно высокорослый горох, от этого высокорослого гороха получилось потомство на ¾ – высокорослое, на ¼ – низкорослое. ІІоследнее навсегда осталось низкорослым. В первом из 3U одна четверть (если ¾ принять за единицу, это будет одна треть) неизменно давала высокорослое потомство: с другими двумя четвертями происходило тоже самое, что случилось с высокорослым горохом происшедшим от первого скрещивания; получалось 3D+1R и дальше повторялась история первых генераций. Отсюда следует, что не во всех высоких экземплярах признак высоты находится в чистоте, часть их еще гибриды; в скрытом состоянии у них еще есть признак низкорослости, который при благоприятных условиях проявится в потомстве.
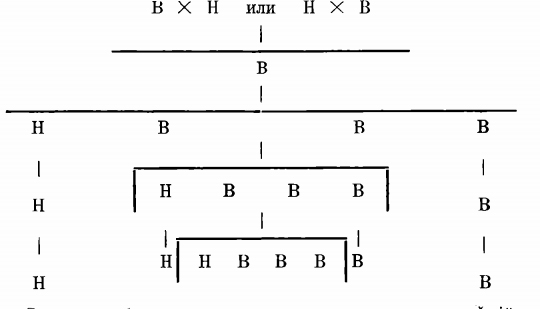
Схема, изображающая потомство четырех поколений, происшедших от скрещивания высокорослого (В) гороха с низкорослым (Н).
Поэтому на самом деле формула для второго поколения (Fs) гораздо сложнее, чем 3D+1R. Четверть этого поколения чисто рецессивная, четверть – чисто доминантная, остальные 2/4 – гибриды. Формула, следовательно, должна быть такова: 1D+1R+2DR, обозначая двумя буквами гибридность потомства. Каждая из этих гибридных четвертей даваля потомство, которое вполне характеризовалось формулою 1D+1R+2DR.
Таковы опыты Менделя. Он проверил их в шести других случаях, особенно под формулу подошли числа которые получились от скрещивания круглозернаго гороха с угловатозерным и желтозерного гороха с зеленозерным. В первом из этих двух случаев пропорция доминантов и рецессивов во втором поколении выразилась так = 2, 96: 1, во втором случае она была 3, 01: І и пропорция доминантов гибридов и доминантов чистых в этом поколении была 1, 93:1 в первом случае и 2, 13: 1 во втором. Опыты Менделя с высокорослым и низкорослым горохом дали цифры более далеко отступающие от его формулы. Из 1064 растений полученных от скрещивания, вышло 787 высокорослых, 277 коротких.
Для того, чтобы проще и яснее представлять опыты Менделя, можно воспользоваться терминами недавно введенными Логансеном. Он называет фенотипичtским – явным – признак или совокупность признаков открывающиеся в индивидууме и он называет генотипическим действительное строение индивидуума с точки зрения его наследственных свойств, т. е. с точки зрения того, какое потомство он может произвести. Так индивидуумы первого поколения (F1) съедобнаго гороха фенотипически были высокорослыми, но генотипически – они гибриды. Точно также треть доминантных индивидуумов второго поколения и фенотипически н генотипически доминантны, так как они производят только чисто доминатное потомство; две другие трети являются доминантными только фенотипически, но генотипически они гибриды.
Согласно опытам Менделя совершенно безразлично при скрещивании, будем ли брать пыльцу доминантнаго растения или пестик его оплодотворять пыльцою растения рецессивнаго, результат будетъ одинъ и тотъ же: все потомство будетъ доминантным. Но в следующем поколении, как мы видели, часть потомства окажется рецессивным. Здесь происходит диссоциация признаков. Закон этой диссоциации и представляетъ собою сущность открытия Менделя. Этот закон открывается во многих случаях, где нет повидимому ни доминантных, ни рецессивных признаков.
В этом отношении поучителен опыт Корренза, производившего скрещивание красноцветной mirabilis Jalappa с белоцветной. Mirabilis jalappa, это – ночная красавица наших садов. Венчик у нея воронкообразный, разрез (она сростнолепестная) пятилепестковый, пять тычинок, родом из южной Америки, корень имеет применение в медицине. У этих разновидностей повидимому нет ни доминирующих, ни рецессивных признаков. От скрещивания по лучились не красные и не белые цветы, а бледнорозовые. Поколение бледнорозовых дало разнообразное потомство. Одна четверть потомства имела цветы белые, все последующие поколения были тоже с белыми цветами; одна четверть потомства имела красные цветы, все его последующие поколения также имели чистокрасный цвет. Очевидно, здесь нетъ гибридизма. Остальные две четверти имели бледнорозовые цветы, и потомство каждой четверти являлось тем же, чем и потомство первого бледнорозоваго поколения. Это потомство нельзя характеризовать формулою ID, 2DR, но путем незначительнаго изменения этой формулы можно получить формулу для случаев вроде того, какой представляет mirabilis jalappa. Обозначим через А признак красноты, через а – признак белизны, через Аа – посредствующий признак (бледнорозовость); тогда потомство, происходящее от Аа, вполне определится формулою: 1А+2Аа+1а26.
Доселе речь шла о судьбе одного признака у скрещиваемых растений. Речь шла о моногибридах. Но Мендель производил опыты и над полигибридами. Он скрещивал разновидность съедобнаго гороха, имевшаго зерна желтые и круглые, с разновидностью, имевшею зерна зеленые и угловатыя. Здесь две пары противоположных признаков: желтый и зеленый, круглый и угловатый.
Чтобы лучше понять результаты опытов Менделя, нужно обратить внимание на то, что зерно съедобнаго гороха, т. е. собственно самый горох состоитъ из внешней оболочки (спермодермы) и из ядра. Это ядро или эмбрион есть ни что иное, как новое растенице образовавшееся из оплодотворенного яйца, оно является уже новым индивидуумом и принадлежит к поколению F1, своим происхождением оно обязано скрещиванию и поэтому является гибридом; напротив, спермодерма происходит из материнской ткани и на образование ее скрещивание не имеет никакого влияния. Для исследования результатов скрещивания нужно иметь в виду лишь эмбрион: желтая или зеленая окраска исследуется в семянодолях, круглая или угловатая форма принадлежит питательным средствам содержащимся в семянодолях.
Первое поколение (F1) представило собою горох, все зерна которого были желтые и круглые. Следовательно, признаки ЖК являются доминантными, ЗУ рецессивными. Мендель высевает эти семена и получает из них. 15 растений, эти растения дают горох, представляющий собою второе поколение (F2). Здесь кроме ЖК и ЗУ являются еще две новыя формы: желтые и угловатые – ЖУ и зеленые и круглые – ЗК. 556 зерен, которые были у Менделя, так распределились по этим четырем формам:
32 ЗУ+Ю8 3K+101 ЖУ+315 ЖК.
Это приблизительно соответствуетъ пропорции:
1 ЗУ+3 ЗК+з3ЖУ+9 ЖК.
Эта формула прежде всего представляетъ собою подтверждение закона диссоциации признаков установленнаго для моногибридов. Если исследовать отдельно каждый из признаков, то окажется, что к полученному потомству вполне прилагается формула 3 D+1B: двенадцать желтых (доминант) и четыре зеленых (рецессив), двенадцать круглых (доминант) и четыре угловатых (рецессив). Но здесь открывается еще и нечто новое. Два признака соединенные в одном родителе: Ж и К, 3 и У, оказывается, во втором поколении могут разделяться и разделяются. Образуются новые соединения признаков, которых не было ни в каком из родителей. Новые соединения образуются в определенной численной пропорции. Если обозначить доминанты через D и D1 и рецессивы через R и R1, то потомство второго поколения определится формулой:
1 RR1+3 RD1+3 R1 D+9DD1.
Таков закон диссоциации в приложении к дигибридам. Но представленная формула, как и аналогичная формула по отношению к моногибридам, только фенотипична: она показывает, какие признаки и в каких комбинациях явились вовне, но она не показывает, что таят в своем существе эти зерна. Для того, чтобы это узнать, нужно от них произвести третье поколение (F3).
Мендель поступил таким образом. Онъ высеял их, разделив их на четыре доли сообразно с их фенотипическою природой. Некоторые из них не взошли, некоторые произвели только безплодные растения, но большая часть произвели плодоносные растения, которые дали зерна представлявшие собою третье поколение.
Вот, что произвел каждый из четырех участков.
Зерна участка ЗУ дали 32 растения, все семена которых также были зелеными и угловатыми. Очевидно, все семена были чисто рецессивными. Так как мы имеем дело с четырьмя признаками ЗЖКУ, то формулой или символом для этих семян может служить обозначение ЗЗУУ.
Зерна участка ЗК дали 102 растения, из них 35 в свою очередь произвели только зерна зеленые и круглые, 67 растений дали зерна исключительно зеленые, но одни из них были круглы, а другие – угловаты. Следовательно, в участке ЗК тридцать пять растений были чисто рецессивными по окраске и чисто доминантными по форме. Формулой или символом их будет ЗЗКК. Шестьдесят семь растений, напротив, были чисто рецессивной расы по окраске, но гибридными по отношению к форме. Их символ – ЗЗКУ.
Зерна участка ЖУ дали 96 растений. Из них 28 произвели зерна искючительно желтые и угловатые, 68 произвели зерна все угловатые, но одни из них были желтые, а другие зеленые. Следовательно 28 растений были чисто доминантной расы по окраске и чисто рецессивной по форме. Символ их ЖЖУУ. Наоборот, 68 были чисто рецессивными по форме, но оставались гибридами по отношению к окраске. Символом их будет – ЖЗУУ.
Зерна участка ЖК произвели 301 растения. Из них 38 дали семена исключительно желтые и круглые. 65 растений дали зерна исключительно круглые, но одни из них были желтые, а другие – зеленые. 60 растений произвели зерна все желтые, но одни из них были круглые, а другие – угловатые. Наконец, 138 растений произвели зерна, из которых одни были желтые и другие зеленые и в каждой из этих категорий были зерна круглые и угловатые. Следовательно из 301 растения участка ЖК были 38 расы чисто доминантной по отношению к обоим признакам – ЖЖКК. Далее 65 были расы чисто доминантной по форме, но гибридной по окраске. Их символ ЗЖКК. Шестьдесят растений представляли собою чисто доминантную расу по окраске, но гибридную по форме. Их символ – ЖЖКУ, Наконец 138 растений являются гибридными по отношению к обоим продуктам. Их формула – ЖЗКУ.
Вывод по отношению к дигибридам может бытъ формулирован таким образом. Если скрещиваются две особи, представляющие две пары отличительных признаков, то первое поколение (F1), будет однообразным, в нем выступят доминантные признаки. Во втором поколении 9/16 потомства будет заключат оба доминирующие признаки; 3/іб будут иметь один доминирующий и один рецессивный признак; 3/16 – другой доминирующий и другой рецессивный; 1/16 будет заключать оба рецессивные признака. Само собою подсказывается, да и Мендель и эмпирически установил и теоретически высказал, что можно чисто математическим путем определить характер следующих поколений, как равно исходя из установленных принципов легко вычислить, какое потомство и в каком поколении явится от скрещивания растений различающихся тремя, четырьмя и т. д. признаками.
Кроме опытов над горохом, Мендель производил опыт над бобами. Эти опыты подкрепили и подтвердили ранее полученные им выводы. Он производил еще опыт над скрещиванием и разведением пчел, но отчет об этих его работах куда-то исчез безследно. Наконец, он производил опыты над ястребинкой, но эти опыты его оказались неудачными. Второе и последующие поколения являлись такими же чистыми, как и первое. Опыты эти производить очень трудно, но Мендель былъ очень искуссный экспериментатор и причина его неуспеха лежала в другом. Дело в том, что ястребинка размножается партеногенетически – девственным путём. У ней можно уничтожить все пыльники, и все-таки явятся семена. Понятно, что такие семена, как и почка, как и отростки, могут только воспроизводить материнское растение.
Но если наследственность у ястребинки являлась для Менделя загадочной, то оставался во всей силе факт, что при скрещивании гороха, бобов и других растений можно предсказывать состав потомства в каком угодно поколении. Факт требовал объяснения. Мендель его дал. Теперь пара взаимно исключающих признаков, как, например, высокорослост и низкорослост, получила согласно предложению Бетсона название аллеломорфов. Когда новая особь развивается из двух гамет, являющихся аллеломорфными, она называется гетерозиготой или гетерозиготной. Когда гаметы одинаковы, она называется гомозиготной. Теорию Менделя часто определяли и определяют, как теорию чистоты гамет. По Менделю гаметы не могут терять своих свойств, но могут их не проявлять. При скрещивании высокорослаго гороха с низкорослым, понятно, все новые особи должны быть гетерозиготными, т. е. образовываться из аллеломорфных гамет. Комбинаций в сущности было только две: соединение высокорослой мужской гаметы с низкорослой женской и соединение низкорослой мужской гаметы с высокорослою женскою. Доминирующим признаком была высокорослость. Все потомство перваго поколения оказалось высокорослым. Это – фенотипически, но не генотипически: низкорослые гаметы находились в каждом растении. Мендель далее не производил скрещиваний. Растения оплодотворялись самостоятельно. В каждом растении было две гаметы, но при оплодотворении растение отдавало новообразующейся особи лишь одну из гамет. Оказываются возможными четыре комбинации. Высокорослая мужская гамета соединяется с высокорослой женской. Низкорослая мужская гамета соединяется с низкорослой женской. Высокорослая мужская гамета соединяется с низкорослой женской. Низкорослая мужская гамета соединяется с высокорослой женской. Теория вероятностей, имеющее болышое применение в опытах Менделя, говорит, что при большом количестве комбинации должны быть приблизительно равными, т. е. что столько же соединится высокорослых с высокорослыми, сколько низкорослых с низкорослыми, сколько мужских высокорослых с женскими низкорослыми, сколько женских низкорослых с мужскими высокорослыми. Опыты Менделя выяснили, что важно только – доминантна гамета или рецессивна, а пол ея не имеет значения. Из этой теории ясно вытекает, что четверть потомства во втором поколении будет низкорослым, три четверти – высокорослыми. Теория предвидит и дальнейшее. Низкорослое потомство во всех последующих поколениях будет низкорослым потому что оно заключает в себе лишь низкорослые гаметы. Потомство образованное исключительно из высокорослых гамет во всех последующих поколениях будет высокорослым. Остаются еще две четверти. Каждая из этих четвертей представляет собою то же, что и первое поколение. В каждой особи имеются аллеломорфные гаметы. При оплодотворении опять возникают четыре комбинации. Одна четверть будет низкорослых, одна четверть чисто высокорослых, две четверти фенотипически явятся высокорослыми, но генотипически заключают в себе и высокорослость и низкорослость и представляют собою то же, что и первое поколение. И от них произойдет такое же потомство, какъ и от первого поколения и так далее до безконечности.
Предыдущее рассуждение относится к моногибридам. Но Мендель дал объяснение явлениям наследственности и у полигибридов. Для этого объяснения он выдвинул принцип расщепления признаков. Если скрещиваются два растения, имеющие две пары аллеломорфных признаков, то все эти признаки оказываются несвязанными один с другим, они могут расщепляться, как угодно, и вступать в какие угодно комбинации. Выше была речь о желтом круглозерном горохе и о зеленом угловатозерном. Мы имеем здесь дело с восемью различными гаметами: желтая мужская, желтая женская, зеленая мужская, зеленая женская, круглозерная мужская, круглозерная женская, угловатозерная мужская, угловатозерная женская. В каждой новообразующейся особи мы должны предположить четыре гаметы: две гаметы цветов и две гаметы формы. Гаметы цветов образуют четыре комбинации: Жм Жж, Зм Зж, Жм Зж, Зм Жж. Маленькими буквами я обозначаю пол. Гаметы формы также обраауют четыре комбинации: Км Кж, Ум Уж, Км Уж, Ум Кж. Соединение гамет цветов с гаметами формы дает шестнадцать комбинаций. Определить по схемам Менделя состав потомства, образующийся из этих комбинаций, не представляет затруднений. Легко также представить себе, как можно прилагать принципы Менделя к растениям, имеющим три четыре и т. д. пар аллеломорфных признаков.
У Менделя получается, что по существу скрещивание не изменяет и не уничтожает никаких свойств организмов. Фенотипически результатом скрещивания является исчезновение тех или иных признаков, но генотипически они сохраняются. Рецессивная гамета может не проявлять себя в целом десятке и более поколений, но встретившись с своей рецессивною подругою, она откроет в новой особи признак, о существовании которого у растения могли уже позабыть. Только если рецессивные гаметы будут упорно не встречаться между собою, они постепенно будут гибнуть безплодно и может произойдти – хотя это трудно допустимо с точки зрения теории вероятностей, что гамета того или иного свойства исчезнет совсем.
2. Теории наследственности развитые независимо от опытов Менделя
Работы Менделя по вопросу о наследственности были сделаны им предметом общественного достояния в 1865 и последующих годах. Но они оставались неизвестными для выдающихся ученых до 1900 г. Чарльз Дарвин в опубликованном им труде – The variations of animals and plants under domesticaton 1868 г. – (в русск. перев. Ковалевскаго: Прирученные животные и возделанные растения) пользуется многими очень сомнительными сообщениями о фактах наследетвенности, но опыты Менделя ему неизвестны. Это неудивительно. Но удивительно, что знаменитый ботаник Негели, бывший в переписке с Менделем, не обратил внимания ученаго мира на его труды. В последние десятилетия XIX столетия возникло несколько теорий наследственности, ставших предметом исследования, проверки и критики, но они не стоят в связи с работами Менделя.
Первою должна быть названа теория Дарвина. Знаменитый биолог, давший теорию происхождения видов, должен был представить объяснение и наследственности и изменчивости организмов. Он и предложил предварительную гипотезу пангенезиса (provisional hypothesis of pangenesis), самым названием показывая, что он еще не убежден в ее истинности.
„Я принимаю, говорит он, что клеточки прежде чем обратятся в совершенно пассивный или окончательный материал, отделяют от себя мелкие зернышки или атомы, которые свободно обращаются по всему телу, и когда снабжены надлежащим питанием, размножаются посредством самоделения, а впоследствии развиваются в клеточки подобные тем, от которых они произошли. Эти зернышки для отличения могут быть названы клеточными почечками (cell gemmulesi, или так как теория клеточек еще не установлена, – просто почечками. Предполагается, что они передаются от рождающих организмов рождаемым, что вообще они развиваются в том поколении, которое непосредственно следует, но часто они передаются в сонном состоянии через несколько поколений и только тогда развиваются. Предполагается, что их развитие зависит от их соединения с другими уже несколько развитыми клеточками или почечками, такими, которые предшествуют им при правильном ходе возрастания. Предполагается, что почечки отделяются каждою клеточкою или вообще единицею не только в зрелом состоянии, но и вообще на всех ступенях развития. Наконец, я предполагаю, что почечки в своем сонном состоянии имеют друг к другу взаимное сродство, приводящее в скопление или в почки, или в половые элементы (семя и яичко). Так, что строго говоря, не эти воспроизводительные элементы и непочки порождают новые организмы, а сами клеточки, находящиеся во всем теле. Вот, предположения составляющие предварительную гипотезу, которую я назвал пангенезисом“. Вот ряд объяснений, даваемых Дарвином на основании этой гипотезы. „Если какое нибудь простейшее первичное животное (protosoa) состоит, как видно под микроскопом, из небольшой массы однороднаго студенистого вещества, то маленький атом, отделившийся гденибудь от нее и питаемый при благоприятных условиях, конечно, воспроизведет все животное; но если верхняя и нижняя поверхности различаются по строению от центральной части, тогда все три части должны отделять зернышки или почечки, которые, соединяясь в силу взаимнаго сродства, и могут составить почки или половые элементы. Точно так, если например, отрезана нога у саламандры, то на ране образуется легкий струп, и предполагается, что под этим струпом неповрежденные клеточки и вообще частицы кости, мускулов, нервов и проч. соединяются с обращающимися по телу почечками тех клеток, которые по порядку следуют за нитаи в цельной ноге; а эти почечки, когда несколько разовьются, соединяются с следуюгцими и т. д., пока не образуется бугорок ткани, почка ноги, а потом и вполне развитая нога». „Дитя, строго говоря, не развивается во взрослаго человека, но заключает βϊ. себе зачатки, которые постепенно и последовательно развиваются и образуют особого взрослого человека. В дитяти, точно так как во взрослом, каждая частица рождает ту же самую частицу, являющуюся в следующем поколении. Наследственность должна быть разсматриваема часто как одна из форм роста, подобная самоделению низкоорганизованнаго одноклеточнаго растения. Атавизм происходит вследствие передачи сонных почечек от предков к потомкам, в которых эти почечки могут иногда развиваться при некоторых благоприятных условиях (известных или неизвестных). Каждое животное или растение можно сравнить с грядкою земли, заключающею в себе множество семян, из которых большая часть быстро прорастает, некоторыя остаются в сонном состоянии, а некоторые погибают. Если иногда говорят, что тот или другой человек содержит в своем теле семена наследственной болезни, то такое выражение очень справедливо понимать в буквальном смысле.
Пангенезис Дарвина прежде всего оказался стоящим в противоречии с фактами. Эмбриология учит нас, что развитие организма, возникающаго всегда из одной клеточки, начинается делением этой клеточки. Это деление клеточки на две совершенно равноправные – факт не только необъяснимый пангенезисом, но стоящий в прямом противоречии с ним. Далее, развитие организма представляет дифференцирование клеточек, размножение их и уразноображение; но теория Дарвина стоит в противоречии с этим фактом, ибо ею предполагается, что зуб образуется из специальных клеточек зуба, и волос из специальных клеточек волоса. Предположение это не верно. Наконец, гипотеза Дарвина ведет к несообразным выводам. „Я вычислил, говорит Негели, сколько клеток, по Дарвину, должно войти в состав липоваго дерева и получил число приблизительно ровное 2000 миллионам. Так как в растительном царстве рост происходит чрез деление клеток, то число клеток, погибших на более ранних стадиях развития, равняется по крайней мере числу их, которые существуют в данный момент. Таким образом, если бы, следуя Дарвину, каждая клетка высылала из себя по атому, то число таких атомов было бы равно 4000 миллионам, по крайней мере. Если же принять, что каждый такой атом есть ничто иное, как молекула белковаго тела с 72 частицами углерода, то и тогда бы величина цветковой крупинки должна бы была быть громадна. Но если быть строго последовательным, то нужно допустить, что от каждой клетки дерева должно быть послано по нескольку атомов, так как клетка сама есть элемент сложный, состоящий из оболочки, ядра и т. п. частей способных изменяться. Кроме того зачатки свойств предков липоваго дерева также должны бы войдти в состав цветени. Таким образом, зародыши атомы Дарвина должны иметь уже свойства чисто метафизические. “
Дарвиновскую гипотезу пангенезиса изменил Брукс (в 1883 г.) Брукс допускает, как и Дарвин, что все клетки многоклеточнаго организма могут отделять крупинки, которые переходят в разные части организма, между прочим, в яйцо, но особенное средство эти крупинки имеют к семенчаткам, в которых они собираются и чрез которые по преимуществу передаются следующим поколениям. Различие от Дарвинской гипотезы (кроме признания первенствующей роли за семенчатками) заключается в том, что, по Бруксу, крупинки не отделяются постоянно от всех клеток, а лишь от некоторых и лить тогда, когда эти клетки попадают в новые условия, нарушающие обычную функцию органа. Стоит какой-нибудь части претерпеть изменение в смысле приспособления к новым изменившимся условиям жизни, и все клеточки этой части тела станут отдавать крупинки, которые при оплодотворении яйца семенем передаются потомству. При этом в оплодотворенном яйце каждая из крупинок, передаваемых семенчаткою, сливается с той частичкой яйцевой протоплазмы, из которой в новом организме должна развиться клетка, соответствующая той, какая в родительском организме отделила данную крупинку. Следовательно, эта вновь происшедшая клетка является ублюдком, помесью двух крупинок – материнской и отцовской, и так как ублюдки вообще расположены к варьированию, то и данные клетки и составленные из них органы будутъ варьировать. Из этих вариаций естественный подбор укрепит некоторые наиболее полезные изменения крупинок, и тогда прекратится отделение крупинок, потому что органы попадут в наиболее благоприятные приспособленные условия, а организация будет передаваться чрез яйцо, и сам избранный естественным подбором организм имеет благоприятные свойства своей организации от яйца, из котораго он произошел.
Гипотеза Брукса, как видим, не объясняет консервативной наследственности, а одну только прогрессивную. Но раз не объяснена первая, то как же объяснить вторую? Гипотеза Брукса, далее, совершенно фантастична и произвольна, как и гипотеза Дарвина.
Эрнест Геккель создал теорию перигенезиса пластидул. Пластидулы – это мельчайшие частицы, из которых строятся пластиды, т. е. клетки и вообще элементарные части организма. Теория пластидул следующая. Весь мир, по миению Геккеля, управляется законами механики, следовательно, этими законами управляются и явления органической жизни. Они будут лишь тогда ясны, когда будут узнаны виды колебаний иластидул, т. е. молекул тела, входящаго в состав протоплазмы клеток. Биогенические процессы подобны периодическим колебаниям волнообразно колеблющихся тел. Ряд предков, расположенных в филогенетическом порядке, походит на волнообразную линию, где индивидуальная жизнь отдельной особи соответствует отдельной волне, а все генеалогическое дерево сходно с разветвляющеюся системой волнообразных линий. Онтогения точно также подобна системе волн, в которой пластиды, т. е. клетки отвечают отдельным волнам. Зная, что клетка есть продукт известным образом колеблющихся молекулъ тела ее составляющаго, мы можем заключить по аналогии, что и их движение должно быть системою ветвящихся волн. Последняя causa efficiens всех биогенических явлений есть характер колебаний пластидул. От этих колебаний зависят все явления наследственности, атавизма и т. д. Негели сурово относится к гипотезе Геккеля. Волны Геккеля, говорит он, суть волны поэтов, но не физиков. Особи биогенических предисов различны и по времени их существования и по материи их составляющей, Волны физика, пробегающие в какой либо среде, постоянно образуются одним и тем же веществом, они отвечают различным колебаниям одной и той же не сходящей с места частички. Физика знает волны пересекающиеся, интерферирующие, но ей не ведомо ни одного случая разветвленной волны. Кроме того Геккель делает и другую ошибку. Пластидула не есть атом, способный колебаться; это есть сложное тело, подобное кристаллической единице. Таким телам едва ли можно приписать колебания подобные колебаниям волн эфира. Самая клетка не есть вещество однородное. Фраза Геккеля, что каждая частичка монеры чувствует так, как и сама монера, слишком смела. Частички плазмы и монера из них составленная так же резко отличаются друг от друга по действию, как частица железа от механизма с колесами и шипами, составленнаго из железа.
Геринг считает наследственность памятью материи. Согласно его воззрению всякой форме, в которой проявляется материи, свойственна память, присущая материи, как одна из ея функций. В силу этой памяти всякий зародыш всегда развивается в определенном направлении и порядке, повторяя все то, что он, так сказать, запомнил от предшествующих поколений. Сродство наследственности с памятью, по Герингу, тем убедительнее, что, как доказано многими фактами, наследственность так же усиливается многократными повторениями, как память развивается упражнением. Известно, например, что чем дальше развивается посредством искусственнаго подбора какая нибудь порода животных или растений, тем легче и легче она получается в последующих поколениях. Как жизненная волна Геккеля, так и память материи Геринга, без сомнения, были бы более уместны, как поэтические метафоры, чем научные термины. Выражения эти дредставляют собою замену понятий словами.
В 1883 г. Вейссман высказал новый взгляд на наследственность и в 1885 г. представил полное развитие своей теории и сделал некоторые дополнения к ней в последующие годы. Наследственность в организмах простых, размножающихся партеногенетически, объясняется Вейссманом, как и некоторыми другими натуралистами, весьма просто. Так как в простейших организмах тело не разделено на органы, а равняется одной клетке, то вещество этой клетки прямо и продолжается в потомстве. Если в течении жизни материнская клетка приобрела какие нибудь изменения, то эти изменения передаются и потомству. Здесь собственно продолжаемость неделимаго (Contuität des Individuums) объясняет явления наследственности. У многоклеточных организмов существует, по Вейссману, продолжаемость зародышевой плазмы (Continuität des Keimplas-mes). Именно, клетки, составляющие тело сложного организма, могут быть разделены на две категории: клетки зародышевые (Keimzellen), служащие для продолжения рода, т. е. половые продукты (яйца и живчики) и клетки соматитические (somatische Zellen), составляющие собственно тело индивида, т. е. все остальные. Строение соматических клеток определяется зародышевой плазмой, сама же зародышевая плазма не изменяется, она растет и переходит от предков к потомкам в неизменном виде. Таким образом оказывается, что, по Вейссману, возможна только консервативная наследственность, а не прогрессивная. Существование последней Вейссман действительно отрицает, он указывает на то, что дети музыканта не наследуют уменья играть на музыкальных инструментах, дети не наследуют уменья читать и т. д. Что касается до постоянно наблюдаемаго сходства между детьми и родителями, то это сходство, по утверждению Вейссмана, заключается не в передаче детям благоприобретенных свойств родителей, а в передаче прирожденных свойств, наприм., цвета волос, глаз, роста, внешняго вида, духовных способностей. Когда Вейссман провозгласил свою теорию, то думали, что он совершенно отрекся от дарвинизма и эволюции, основывающихся прежде всего на прогрессивной наследственности; но Вейссман в последующих статьях заявил, что он признает эволюционную теорию происхождения видов и представил объяснение того, как примиряется его гипотеза наследственности с эволюционной теорией. Bo 1) он допустил, что зародышевая плазма может изменяться, но только изменяться весьма незначительно под влиянием внешних условий, во 2) он указал источник изменений плазмы в половом размножении. В последнем смешиваются две плазмы – две наследственные генденции. Отсюда окончательное строение плазмы непременно должно быть измененным сравнительно с первоначальным, отсюда изменяемость видов. Ключ к объяснению происхождения видов Вейссман таким образом видит в комбинациях при двусторонней наследственной передаче. Вейссману возражал Вирхов, указывавший на наследственность болезней как на опровергающую теорию Вейссмана. Но Вейссман попытался объяснить эту наследстоенность инфекционным характером этих болезней; он говорил, что причина их можег заключаться в особых бацилах, что эти бацилы передаются от родителей к детям не в свойствах зародыша, а прямо переходят с одного организма в другой.
Об этой теории Вейссмана должно сказать то же, что и о других теориях. Bo 1) она произвольна. Вейссман не мог, конечно, наблюдать собственно зародышевой плазмы, ее неизменяемости, ее молекулярнаго строения, он даже не может доказать какими нибудь побочными данным, что таковая существует. Во 2) несомненно, что часто передаются наследственно болезни, не имеющие инфекционного характера, какова, например, эпилепсия. Что источник этой болезни лежит не в бациллах, это доказывается опытами Броуна Секара и Вестфаля над морскими свинками. ІІоследними ударами молота по голове вызывал в них искуственно эпилепсию. Очевидно, что бациллы здесь не при чем. Этот опыт губителен для теории Вейсемана.
Выступавший против Дарвина Келликер прннял, что существует общий закон развития органического мира, закон, подобный тому, по которому развиваются отдельные организмы. При чем, по Келликеру, развитие органическаго мира может совершаться и действительно совершается скачками, а не в строгой постепенности. Изменения в организмах, говорит Келликер, состоят во 1) в изменении существующих органов, во 2) в возникновении новых морфологических единиц. Изменение существующих органов – Келликер допускает – может происходить под влиянием внешних условий, но возникновение новых органов имеет свою причину в законе общаго развития, а не в случае. Странно допустить влияние подбора на возникновение новых органов, когда эти органы вначале будут только лишним или даже вредным бременем для организма. Появление спинной струны у первых млекопитающих, первых позвонков, первых кровеносных сосудов, – все это непонятно с точки зрения естественнаго подбора. Затем, если обратить внимание на то, что почти все более или менее крупные изменения зародыша и прежде всего все настоящие новообразования органов случаются в самые ранние поры зародышевой жизни, так у куринаго зародыша все главные органы закладываются в первые 4–5 дней его существования, у человека к концу первого месяца его утробной жизни; если обратить внимание на это обстоятельство, то очевидно нельзя представить себе развитие организмов таким образом, что они сначала проходили все стадии безпозвоночных животных, а потом стали принимать признаки позвоночных. Развитие нужно представлять иным образом. Пo Келликеру, возможны и действительно бывают такие случаи, что от родителей, стоящих на низших ступенях, положим, кишечнополостных или червей прямо возникнут насекомые. Для обоснования и разъяснения своей теории он обращается к явлениям метагенезиса (перемене генераций)27, гетерогонии28 и педогенезиса29.
Простейшую форму метагенезиса представляют некоторые кольчатые черви (Syllis). Они раздельнополы и имеют вполне развитые, хотя и весьма просто устроенные, половые органы. Из оплодотвореннаго яйца возникает личинка, а из нее молодой червь, проходя типические стадии развития. Но почти взрослый сложившийся червь, не имеющий только вполне развитого полового аппарата, может делиться поперек на несколько частей, от которых каждая образует нового маленького червя. Эти маленкие черви представляют, следовательно, собою поколение, происшедшее путем безполого размножения, путем деления материнского организма. Они растут, достигают половой зрелости и размножаются посредством оплодотворенных яиц, производя следующее поколение путем полового размножения как произошел и первый червь. Более сложное явление метагенезиса наблюдают у некоторых медуз. Эти морские животные имеют обыкновенно вид студенистаго полупрозрачного колокола и размножаются посредством оплодотворенных яиц. Из яйца развивается овальная личинка, плавающая некоторое время в море и, наконец, укрепляющаяся одним своим концом на каком-либо подводном предмете. На свободном конце ее тела появляется ротовое отверстие, а вокруг него выростают щупалыда. В этом виде превращенная личинка медузы называется сцифистомою и напоминает собою сидячий полип вроде пресноводной гидры, водящейся в наших прудах. Сцифистома растет в вышину и начинает подразделяться поперечными перехватами на части, а ее щупальцы претерпевают обратное развитие, т. е. уничтожаются. В этой стадии развития животное называется стробилою. По краям сегментов стробилы образуются затем новые отростки (щупальцы) и мало по малу сегменты отделяются в виде плоских дисков с отростками, которые плавают отдельно и превращаются постепенно в взрослых колоколообразных медуз. Здесь, таким образом, половые и безполые поколения оказываются совсем непохожи одно на другое, здесь являются превращения. Теперь обратим внимание на то, что сцифистома в известной стадии развития весьма напоминает гидру. Исходя из этого, Келликер предполагает, что возможно, что некогда гидроподобные организмы размножаясь, произвели дотоле несуществовавшие формы медуз.
Явления метагенеза указывают по Келликеру на возможность происхождения совершенно новых существ впезапно, т. е. без всяких постепенных переходов. Возможность этой внезапности открывается также из фактов гетерогонии. Под именем гетерогонии разумеют те случаи, когда одно ито- же животное при размножении своем является в двух или более различных формах зрелых в половом отношении, Так, наприм., существует маленькая круглая глиста, аскарида лягушачья (ascaris nigrovenosa), которая живет в двух различных формах – в легких лягушки и во влажной земле. В легких лягушки живет гермофродитическая форма, яйца которой развиваются из раздельнополых глист, имеющих признаки рода Rhobditis. Эти молодые робдитисы из легких лягушки нереходят в ее пищеприемное горло и пройдя чрез пищеварительный канал, с испражнениями попадают на землю, после чего живут в земле свободно, пока снова не попадут в рот и легкие лягушки, где они производят первоначальное гермофродитическое поколение. Так, оказывается, что из одного и того же начала возникают ascaris nigro-venosa и rhobditis nigrovenosa.
Что касается до педогенеза, то Келликер цитирует открытые пр. Вагнера о размножении личинок мух, как подтверждающее его теорию разнороднаго происхождения. Вагнер нашел, что личинки некоторых мух (cecidomyi-dae) могут производить внутри своего тела новое поколение подобных же личинок. Эти последние поедают внутренности матери и выходят наружу, после окукливаются или же вторично производят внутри себя поколение личинок, которые уже подвергаются окукливанию. Из куколок развиваются мухи. Можно допустить, говорит Келликер, что личинки насекомых существовали некогда, как самостоятельные формы и внезапно стали производить не подобных себе личинок, а новые существа, т. е. современных мух.
В подтверждение теории Келликера о возможности внезапных переходов организмов одного вида к организмам другого вида можно указать еще на наблюдения и опыты Уоллеса и Вейссмана над бабочками. Существуют, наприм., две формы европейских бабочек vanessa levana и vanessa prorsa, резко различающиеся между собою цветорасписаетам. Первая – буро-желтаго цвета с черными пятнами и полосками, вторая – черная с широкой белой перевязью на обоих крыльях. Эти две формы долгое время считались за два различных вида, но впоследствии оказалось, что различие окраски обусловливается влиянием времен года: vanessa levana есть перезимовавшая в куколке vanessa prorsa. Теперь существуют самостоятельно и prorsa и Іѳуапа. Вейссман развил теорію, что prorsa произошла от levana во время ледниковой эпохи. С точки зрения Келликера, находящей в данном случае опору в фактах настоящаго, такой переход мог произойдти внезапно.
Теория Келликера отрицает знаменитое древнее положение: natura non facit saltum, положение, надо заметить, представляющее собою такой же бэконовский idolon для современных натуралистов, каким для прежних являлся horror vacui в природе. Теория эта, далее, устанавливает новый взгляд на отношение организмов между собою. По Дарвину и другим, различные организмы тем ближе генетически, чем они ближе физически, по Келликеру этого не оказывается. Непосредственным детищем червеобразного существа может быть musca domestica – организм совсем другого типа, стоящий рядом с musca caesar, vomo-toria и могущий не имет с ними никакого родства. Келликер идет даже далее, он утверждает, что организмы одного вида могли происходить от совершенно различных предков, почему и называет свою гипотезу полифилитическою в противоположность дарвиновской, которая, как он законно говорит, непременно должна быть монофилитической. С точки зрения Келликера, значит, человек мог родиться – даже и в библейском смысле потенциально совершенным – от какого нибудь вовсе непохожаго на него млекопитающаго, но мог, конечно, родиться и от обезьяны.
В 1884 году появилась теория происхождения видов, – Негели (Mechanisch= physiologische Theorie der Abstammungslehre. München und Leipzig 1884) приближающаяся в некоторых пунктах довольно близко к теории Келликера. Как и Келликер, Негели в борьбе за существование и естественном подборе не видит достаточных факторов для образования новых видов. Они по его воззрению, только устраняют с жизненной арены менее приспособленные организмы и таким образом производят пробелы (Lucken) между видами, но не создают новых видов. Появление таковых отчасти должно быть приписано влиянию внешних условий, но главным образом оно должно быть приписано принципу совершеяствования (Vervolkommengsprincip). Этот принцип механическаго свойства и есть ничто иное, как закон инерции в области органического развития. Раз эволюционное движение началось, оно не может остановиться, а должно продолжаться в определенном направлении. Негели замечает, что если бы дарвиновские факторы заправляли развитием организмов, то изменения последних происходили бы во всевозможных направлениях, и в действительности не было бы того генеалогического дерева организмов, которое представляет нам окружающий мир. Изложим теорию Негели. По воззрению Негели, составившемуся на основании изучения строения крахмальных зерен и клеточных оболочек растений, все организованные тела состоят из мельчайших частиц, большею частию правильной кристаллической формы, так называемых мицелл (объем одного мицелла, по Негели, равенъ 0,0000000021 кубич. микромиллиметров, принимая микромиллиметр равнымъ 0,001 миллиметра). Понятно, что такие мицеллы не видны ни в какие микроскопы, тем не менее мицеллы не представляют собою нечто простое, нет, каждая из них представляет определенную группу или систему молекул. В сухом состоянии тела мицеллы могут плотно прилегать друг к другу; но в присутствии воды они раздвигаются на известное расстояние, так как их взаимное притяжение не столь сильно, как их притяжение к воде. Таким образом каждый мицелл одевается тонкой водяной оболочкой. Всякая клеточка состоит из таких мицелл, составляющих ея протоплазму, ядро и оболочку. Яйца, из которых развиваются организмы, понятно, как и самые организмы, состоят тоже из мицелл. Не все яйцо представляет собою начало, из которого развивается организм, болыпая часть яйца, как известно, состоиг из питательнаго материала для остальной части – зародыша в собственном смысле, который Негели называет идиоплазмой. Все признаки, отличающие взрослые организмы друг от друга, содержатся уже с самаго начала развития в строении идиоплазмы, каждый организм обязан своим возникновением особой модификации этого вещества. Таким образом, идиоплазма как бы заключает в себе множество зачатков будущих свойств организма. Чрезвычайная сложность строения, которая должна быть приписана идиоплазме высших животных и растений, является результатом всего племенного развития. У низко организованных предков их она была весьма проста. Ее можно сравнить, говорит Негели, с недисциплинированным средневековым войском, так как сложная идиоплазма уподобляется хорошо обученной армии с подразделениями и с подчинением частей. По воззрению Негели идиоплазма представляет собою как бы сеть, распространенную по всему организму и связуюшую все его части между собою. Эта сеть проходит через все клетки, нити ее особенно сосредоточены в ядрах клеток. Негели думает, что сетеобразное расположение протоплазмы и ядернаго вещества в клетках растений находится в соответствии с разветвлением идиоплазмы по телу. Шнуры этой идиоплазматической сети и состоят из параллельных рядов мицелл. Мицеллы соседних рядов динамически связаны друг с другом так, что возбуждение, испытываемое каким-либо одним рядом передается и на соседние ряды (подобно элекрической цепи). При всяких изменениях идиоплазмы, влекущих за собою появление новых наследственных свойств, все шнуры ее сети, все ряды ее мицелл претерпевают изменения. Эти изменения могут происходить или материальным или динамическим путем. В первом случае при изменении идиоплазмы в какой-либо части тела от этого места распространяется перестройка посредством двигания новых мицелл во все части организма чрез тончайшие отростки протоплазмы, соединяющие отдельные клеточки между собою. Если же имеет место второй способ, то значит, что при каждом изменении в напряжении мицелл в одном месте изменяется тотчас относительное напряжение их и во всем организме. Какой именно способ изменения мицелл имеет место в действительности, Негели не решает. Причины изменения в мицеллах двояки: одни действуют постоянно, это те, которые вызывают рост мицелл. Между существующими мицеллами выдвигаются новые, образуются новые ряды и т. д. Хотя при этом собственно происходит рост только организма, однако, и здесь уже в мицеллах происходят, правда, очень незначительные, но все таки такие изменения, которые изменяют организацию потомков этого организма. Так как внедрение новых мицелл между уже существующими обусловливается природой и расположением последующих, то из этой геометрической конфигурации может возникнуть только новая нетожественная с нею конфигурация. „По принципам механики, говорит Негели, если раз состояние превратилось в а, то состояние b не может уже опять превратиться в а, но лишь в следующее с и т. д.“. Другие причины – случайные, производящие внезапное изменение структуры мицелл. Последним причинам Негели не придает важнаго значения.
Установивъ причины и законы изменения организмов Негели происхождение видов представляет себе таким образом: сначала образовался, да и теперь образуется из неорганической материи белок, молекулы которого взаимным притяжением образовали мицеллы. Между мицеллами начали вдвигаться новые мицеллы. Наконец их соединилось столь много, что они не могли сдерживаться взаимным притяжением и они распались на несколько групп, образовав первичные организмы с безпорядочной молекулярной структурой (пробии – προβιοι). Затем, явились более сложные организмы, мицеллы которых находились уже в более упорядоченном состоянии и соотношении и т. д. При этом нужно заметить, что, хотя идиоплазма изменяется в организмах постоянно, эти изменения могут долго не обнаруживаться во внешних изменениях организма, и затем через много поколений могут выразиться сразу решительным образом. Таким образом Негели, как и Келликер, признает скачки. Человек по теории Негели, как и по Келликеру, мог произойдти независимо от обезьяны и, от каких бы то ни было животных, особым путем, путемъ совершенно самостоятельным от особаго белка и пробия. Родственников человека не существует в современном животном царстве, как и вообще все животные виды, можетъ быть, не родственны между собой, и каждый имеет свой особый корень.
В то время, как на европейском материке ученые, исходя из сознания неудовлетворительности теории Дарвина, выступили с собственными теориями, соотечественник Дарвина Роменс представил теорию физиологическаго подбора для объяснения происхождения видов, теорию, которая восполняла учение Дарвина и лишала силы многие из направленных против него возражений. Против теории Дарвина прежде всего были приведены три следующие возражения: 1) безплодность при скрещивании видов, 2) растворение изменений организмов через скрещивание и 3) безполезность незначительных особенностей, приобретенных какими нибудь организмами и вследствие этого потеря, а. конечно, не развитие этих особенностей; эти три факта, утверждали, устраняют возможность образования новых видов чрез естественный подбор и борьбу за существование. Роменс, пытаясь спасти теорию Дарвина, изменяет его учение таким образом. По его мнению в ряду изменений, которые возникают в организмах, для образования новых видов имеют существенное значение изменения в половой системе (воспроизводительные). Изменения эти действительно бывают и состоят в том, что индивидуум оказывается безплодным при скрещивании с одним и плодовитым при скрещивании с другими индивидуумами. Этим фактом устраняется нужда в миграции, которую Морис Вагнер признал необходимою для того, чтобы естественный подбор производил те следствия, которые ему приписывал Дарвин. Этим фактом некоторые организмы обособляются от других организмов им подобных и живущих в той же местности. Эти некоторые организмы, приобретая какиея-либо особенности, очевидно, не будут терять их чрез скрещивание. Особенности будут унаследоваться, и так, наконец, образуется новый вид. ІІримеры изменения в воспроизводительной силе лучше всего показать на цветах. Представим себе, что какой нибудь цветок приобрел или получил такую особенность, что его цветочная пыль созревает двумя неделями позже, чемъ пыль других цветов того же вида, представим, что таких цветов явилось несколько, очевидно, скрещивание будет возможно только между ними, а не между всеми цветами этого рода. Отсюда, если эти запоздалые цветы приобретут какую-нибудь особенность, то эта особенность передастся их потомкам. С точки зрения Роменса физиологический подбор должен был одним из главных факторов, содействовавших происхождению человека от обезьяноподобнаго предка.
Рассмотрим теперь критически эти десцендентные учения. Во всех этих трех учениях есть одна общая особенность: теории эти не имеют под собою фактов, можно даже сказать больше, теории эти, вообще, не имеют за собою оснований. Первые две из них весьма близки между собою по своему метафизическому характеру. Современные ученые очень презрительно относятся к метафизике и, однако, очень охотно прибегают к ее помощи при составлении гипотез. Негели хочет объяснить вес органический мир из комбинации мицелл – этих своего рода органических атомов. Причина всего, по воззрению Негели, лежит только в перемещении мицелл. Келликер не дал теории идиоплазмы, но очевидно, что мицеллы Негели разъясняют и его теорию, иначе его теория явллется не только не обоснованной, но и не выясненной. Но во 1) существуют ли эти мицеллы? Во 2) если они существуют, то каким образом они приобретают свойства растительных организмов? Негели говорит об их динамическом состоянии, поясняет аналогиями с тем, что нам неизвестно, так как объяснения элекрических явлений, на которые ссылается Негели, ведь, не имеется, как вообще не имеется, что может засвидетельствовать каждый физик, удовлетворительной теории электричества. В 3) каким образом комбинации мицелл получили способность ощущения? Негели не отвечает удовлетворительно ни на один из предложенных вопросов, как не мог ответить на те же вопросы и Демокрит, предлагавший вместо мицелл Негели атомы огненной природы. А между тем уже многими учеными представлены в явлениях царства растительного многие серьезные возражения против теории мицелл. Первым начал возрджать Шимпер. Негели построил теорию мицелл на основании наблюдений над крахмальными зернами, будто бы увеличивающимися в объеме чрез интуссусцепцию, т. е. значит растущими сразу во всех своих частях, но Шимпер показал, что рост крахмальных зерен совершается не чрез интуссусцепцию, а чрез наложение (все равно, как увеличивается объем дерева), это совершенно разбивает мицеллярную теорию. Но теория Негели страдает еще и другими недостатками. Во 1) она не объясняет происхождения полезных изменений в организме; во 2) – и это самое главное – она стоит в противоречии с фактами регрессов в природе. По Негели, организмы должны постоянно усовершенствоваться, усложняться, но есть много организмов, не изменившихся в течении многих геологических веков, головоногие моллюски из рода nautilus существуют с силурийской эпохи до наших дней. Далее, существуют организмы, организация которых упростилась в настоящее время сравнительно с тем, какою она была в прошедшем, таковы киви, потерявшие крылья, таковы змеи, потерявшие ноги. Эмбриология асцидии дает основания полагать, что они прежде стояли выше, чем теперь Асцидии организованы довольно низко и сидят неподвижно, приросшие к камням, устилающим дно моря вблизи берегов. Вся жизнь их состоит в глотании и продеживании морской воды с разными мелкими организмами и в размножении. Но насколько однообразна, невзрачна организация и жизнь взрослых асцидии, настолько же интересна их история развития. Первые стадии развития лица до такой степени похожи на соответственные фазы развития позвоночного животного – ланцетика, что их невозможно различить. Из яйца далее вылупляется свободно плавающая личинка с подвижным хвостом, с внутренним скелетом (с спинною струною – chorda dorsa· lis), с довольно развитою нервною системою и с одним глазом. Но вскоре эта довольно высоко организованная личинка прикрепляется к какому – нибудь камню и начинает терять одну черту совершенства за другою. Хвост исчезает, внутренний скелет пропадает, нервная система упрощается, глаз теряется, мало-по-малу животное обростает оболочкой и превращается в взрослую асцидию, организация которой гораздо ниже организации личинки. Согласно установившимся воззрениям эволюционистов эта история развития асцидий указывает на то, что они произошли от более совершенных предков, чемъ они сами. Теория Негели не может допустить таких фактов, но современное естествознание считает их несомненными. Очевидно, должно поэтому признать несомненной и несостоятельность теории Негели.
Келликер кроме ничего недоказывающих аналогий не представил никаких оснований в пользу своей теории. Этими основаниями для его теории в значительной мере могли служить соображения Негели, но раз эти соображения оказываются не имеющими значения, то и теория Келликера является не имеющею никакой опоры.
Теория Роменса, претендующая дополнить учение Дарвина, в сущности не делает это учение более правдоподобным. Естественный подбор обусловливается, по Роменсу, физиологическим подбором. Те индивидуумы, от которых имеет произойти новый вид, прежде всего теряют плодовитость при скрещивании с индивидуумами стараго вида. Но что означает собою эта потеря плодовитости? Без сомнения, некоторое болезненное изменение в организме. Организм вполне нормальный при скрещивании с каждым вполне нормальным организмом должен дать плод. Затем, это болезненное изменение может, ведь, не сопровождаться другими изменениями, как, напротив, очень часто бывает и тому свидетельство безчисленные разновидности, что совершенно различные изменения, возникают в организмах одного и того же вида, нисколько не препятствуют плодовитости этих разновидностей при скрещивании. Теория Роменса предполагаег обособление некоторых индивидуумов, затем, возникновение в их среде изменений, распространяющихся на всю среду. Но здесь прежде всего возникает вот какое затруднение. Обособление индивидуумов в одном поколении должно ли продолжаться на следующие поколения, начало консервативной наследственности не возьмет ли верх, и потомство обособленных организмов не сделается ли снова способным к скрещиванию и плодовитым со всеми организмами своего вида? Наконец, допустим окончательное обособление организмов, но, ведь, нет никаких причин к тому, чтобы изменения в организации одинаково происходили во всех их потомках. Если несколько особей, получивших какую либо особенность, стали плодовитыми только между собою, то положим, есть вероятность, что они сохранят эту особенность (заметим при этом, что здесь потеря плодовитости и возникновение особенности являются внезапными, так что здесь собственно нет согласия с дарвинизмом, а с учениями Келликера и Негели), но каким же образом эти особи и их потомки станут выделяться в особый вид, ведь, у одних из них может явиться какая либо новая особенность, а у других нет, и скрещивание будет действовать на них нивеллирующим образом. Допустим, что под влиянием каких либо условий в 2 из 20 обособленных физиологическим подбором особях явились какие либо особенности, положим, голубой цвет глаз у кошек; при скрещивании этих двух с остальными 20 не должен ли этот признак раствориться и исчезнуть. Таким образом, скрещивание, правда, ограниченное не устраняется здесь совсем и приводит к тем же роковым последствиям для образования нового вида, как и собственно в дарвиновской теории. Особенно, если принять во внимание размножение организмов в геометрической прогрессии: от 20 происходит 200, от 200–2000, причем большннство не имеет новых признаков; очевидно, особенности меньшинства должны исчезнуть пред давлением болышинства.
Но самою лучшею критикою изложеных теорий являются опыты Менделя. Законы числ установленные Менделем совершенно несогласимы с метафизическими снекуляциями предложенных учений.
Кроме того, что предлагались теории наследственности, учеными делались попытки установить законы наследственности. Мне думается, что такую попытку представил уже Кетле (1796–1874) своею теориею фиктивнаго средняго человека. Наши статистические рассчеты обыкновенно имеют в виду никогда не существовавшаго средняго человека – средний рост, средние способности, средний аппетит, средние силы и т. д. Действительные люди представляют собою по своим свойствам большее или меньше уклонение от средняго типа. Но законом жизни является то, что человечество неизменно возвращается к этому среднему типу, все, что далеко уклоняется от него, погибает.
За всем тем основателем статистическаго метода изучения явлений наследетвенности является не Кетле, а Френснс Гюьтон, двоюродный брат Чарльза Дарвина. Результаты своих исследований он изложил в книге Hereditary genius, its laws and consequences. Порусски она переведена под заглавием – „Наследственность таланта. 1875“. Гальтон изучал такс и людей. Он составил очень много генеалогических дерев, отмечая физические, умственные н нравственныя свойства всех членов семьи. Быт создан специальный орган Biometrica, в котором сообщалось о результатах исследований над явлениями наследсгвенности. Создана специальная лаборатория Eugenic’s Labora-tory. Исследователи задались целью определить законы передачи благородных свойств путем наследственности. Сам Гальтон попытался установить такой закон. Если совокупность свойств особи обозначить единицей, то половина этих свойств получена непосредственно от родителей – пo одной четверти от каждаго; четверть свойств получено от двух дедов и двух бабушек – по одной шестнадцатой от каждаго, одна восьмая свойств от прадедов и прабабушек – по одной шестьдесятъ четвертой от каждаго и каждой. Таким образом свойства особи представляютъ собою сумму безконечно нисходящей геометрической прогрессии, нисходящие члены которой соответствуютъ восходящим поколениям предков:
½, ¼, 1/8, 1/16, 1/32 и т.д.
А наследие от каждаго из предков в восходящем порядке выразится прогрессией:
¼, 1/16, 1/64, 1/256, 1/1024 и т.д.
Гальтон и Мендель родились в одном и том же 1822 году. Оба они занялись изучением явлений наследственности. Гальтон быстро приобрел себе широкую известность. Мендель приобретает известность только теперь. Но славе Гальтона придется померкнуть в лучах славы Менделя. Работы Гальтона практически безплодны, теоретически сомнительны. Ученик и продолжатель дела Гальтона Пирсон (Pearson) счел нужным изменить его математические формулы и еще усложнил их. Самая сложность формул делает их практнчески неприложными, но главное эти формулы дают нам все какие то средние величины неприложимые к отдельным случаям и не дающие возможности предвидеть ни одного факта. Монах Мендель своим опытами и формулами удовлетворил строгому положению отца позитивной философии Конта – savoir, c’est ргёуоіг, знать значит предвидеть.
3. Опыты и гипотезы менделистов
Опыты и гипотезы Менделя, после тридцатипятилетней безызвестности ставшие всемирно известными, задали науке множество вопросов, заранее определяя метод их исследования и отчасти подсказывая их решение. Многие ученые пошли по пути указанному брюнским монахом, образовалось движение, получившее имя менделизма, и примкнувшие к нему получили прозвание менделистов.
Виктор Грегуар в обстоятельной работе о Менделе и менделистах назызает следующих биологов, занимающихся проблемами менделизма. В Англии – Бетсон и его сотрудники: Зондерс, Пеннетт, Дюрхем, Герст, Грегорп, Локк, Уэльдел, Донкастер. В Германии – Корренз, Баур, Геккер. В Голландии – де-Фриз, Тамхиес. В Франции – Кено, Кутан, Вильморен. В Австрии – Чермак. В Швеции – Нильссон – Эле. В Швейцарии – Ланг. В Соединенных Штатах – Давенпорт, Пёлл, Мак Дугал, Морган, Кестл, Ист, Гагедорн, Тоуэр. В Японии – Ту-айяма. Из специальных монографий посвященных ме-делизму (кроме упомянутыъ мною выше) Грегуар называет: Бетсона (Bateson) – Mendel’s principies of Heredity. 1909; Корренза (Korrens) – Ueber Vererbungsgesetze. 1905; Баура (Baur) – Einführung in die experimentelle Vererbungslehre. 1911; Гольдшмидта (Goldschmidt) – Einführung in die Vererbungswis senschaft. 1911; Геккера (Haecker) – Allgemeine Vererbungslehre. 1911. Грегуар перечисляет следующие периодические издания специально посвященные менделизму: Reports to the Evolution committee of the Royal Society (Лондонъ). Publications oi the Carnegie Institution (Вашингтон). Zeitschrift für induktive Abstammungs – mnd Vererbungslehre (Берлин), American Naturalist (Нью йорк), Journal ot Genetics (Лондон). Ho кроме журналов названных Грегуаром можно указать еще периодические издания не посвященные специально менделизму, но отводящие ему значительное место. Таково Revue des Questions scientifiques, где помещена работа самого Грегуара. Таково всемирно распространенное Revue Scientifique, где на пространстве последних годов дан ряд статей о мен делизме (Гадесо – Le Mendélisme, 23 декабря 19.11; Лютцъ – La Génétique, 6 января 1912; Бларингем – Les Problèmes de l’Heredite, 24 февраля 1912; Бларингемъ – Les Problèmes de Biologie appliquée, 2 марта 1912; Аперт – Les Problèmes de l’Hérédité, 12 іюля 1913).
Ha русском языке, не считая переводных работ, о менделизме писалось немного. Проф. Тимирязев неоднократно вспоминал о нем на страницах „Вестника Европы“. В одной статье он сказал, что опытам Менделя придают преувеличенное значение, потому что он монах. В другой статье он признал, что Мендель дал хотя единственное и неболышое, но дельное исследование, однако закончил статью заявлением, что движение менделистов (он называет их мендельянцами) представляет собою церковную реакцию. Такой взгляд трудно понять, да едва ли и нужно стараться его понять. Появлялись еще в журналах библиографические заметки о менделизме. В Русскаго Богатства 1913 г. Филипченко в статье „проблема пола в современной биологии“ показывает, как теориею Менделя можно объяснить тот факт, что мужские и женскиея особи рождаются приблизительно в равном количестве.
Менделисты занимаются главным образом развитием положений Менделя. Стремятся они дать полную и точную терминологию для явлений и фактов, но к сожалению разные авторы выступаютъ с различною терминологиею, и читателям приходится усвоять несколько номенклатур, что и нелегко и безполезно. Так гомозиготы другими авторами называются гомодинамными, гетерозиготы называются гетеродинамными. Факторы наследственности называются генами, детерминантами. Менделисты пытаются вводить и некоторые изменения в положение Менделя. Такова предложенная ими, но принятая не всеми теория присутствия; отсутствие факторов. Мендель для каждой пары аллеломорфных признаков предполагает пару факторов: фактор высокорослости и фактор низкорослости, фактор окрашивающий в желтый цвет, и фактор окрашивающий в зеленый цвет. Теперь пытаются представить дело иначе. Доминантный признак производится некоторым специальным фактором, но рецессивнаго фактора не предполагают. Согласно этому взгляду высокорослый горох есть тоже, что и низкорослый горох, но кроме того, что есть в низкорослом горохе, в высокорослом горохе есть фактор сообщающий ему высокорослость. В низкорослом горохе никакого специальнаго фактора низкорослости нет. Когда при оплодотворении в новообразующуюся особь входит фактор высокорослости, особь будет высокорослой; когда этот фактор не войдет, особь будет низкорослой. Схема потомства здесь будетъ та же, что и у Менделя. Несколько изменено только объяснение.
Прилагают это объяснение и к тому случаю, когда от скрещивания получаются формы промежуточные между скрещиваемыми. От скрещивания красной mirabilis Jalappa – ночной красавицы с белою получается бледнорозовое потомство. Объясняют это так. В красной mirabilis jalappa фактор окраски присутствует в двойной дозе. В белых цветах никакого наследственнаго фактора нет. При скрещивании фактор окраски переходит в новообразующуюся особь в одной дозе, потому что этот фактор дается лишь одним из родителей. Вследствие этого потомство и является с бледнорозовой окраской. Но в следующем поколении фактор окраски передается и в двойной дозе (четверти потомства) и в одной дозе (двум четвертям) и не передается совсем (одной четверти). Опять процесс совершается вполне по схеме Менделя.
Менделисты чрезвычайно расшили территорию опытов и наблюдений. Они установили очень много новых фактов. Порою эти факты на первый взгляд представляются стоящими в несогласии с положениямит Менделя, но потом оказывается, что ими только подтверждаются эти положения. Было произведено множество опытов скрещивания мышей. Скрещивали серых домашних мышей с альбиносами, т. е. с белыми мышами. Серый цвет оказывался доминирующим. Во многих случаях дело шло прямо по схеме Менделя. Первое поколение оказывалось все серым, во втором поколении четверть потомства была белою и производило исключительно белые поколения. Но в других случаях оказалось нечто иное. Первое поколение от скрещивания серых мышей с альбиносами было исключительно серым, но во втором поколении часть мышей оказалась сераго, часть – белаго и некоторая часть – чернаго цвета. Пропорция была такова: 9 серых, 8 черных, 4 белых. Нетрудно видеть, что эта формула в сущности таже, которую Мендель предложил для случаев дигибридизма – двух пар аллеломорфных признаков. Значитъ у мышей подвергавшихся скрещиванию имелось не по одному противоположному признаку, а по каким-то двум. Здесь имелось совместное действие двух факторов, принадлежащих к двум парам аллеломорфных признаков.
Кено, не принимающий гипотезы „присутствия“ и „отсутствия“ дал такое объяснение факту. Он допускает, что существует фактор – хромоген, который обусловливает возможность окраски шерсти, но сам не дает никакой окраски. Затем имеется другой фактор, который своим действием на хромоген дает пигментацию, т. е. окраску, но который в отсутствии хромогена не может произвести пигментаций. Хромогенный фактор (X) является доминантным по отношению к альбинизму (А). С другой стороны фактор, дающий серую окраску (С), является доминантным по отношению к фактору, дающему черную окраску (Ч). Кено для серой мыши дает формулу ХС, т. е. хромоген + серая окраска, для белой мыши даетъ формулу A4, т.е. альбинизм)-черная окраска, не проявляющая себя вследствие отсутствия хромогена: скрещивание серых мышей с белыми дает потомство, имеющее формулою ХАСЧ. Очевидно, это потомство будетъ серым. Это потомство дает четыре сорта гамет для каждаго пола ХС, СЧ, АС, – A4. Следующее поколение согласно Менделю должно соответствовать представляемой схеме.
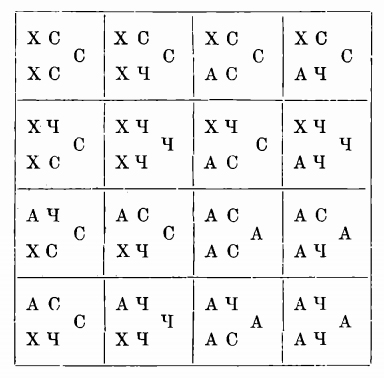
Везде, где хромоген соединится с серой окраской, потомство будет серым. Где хромоген при отсутствии серой окраски соединится с черной, потомство будет черным. Везде, где хромоген отсутствует, потомотво будетъ альбиносным. Из таблицы видно, что на 16 особей, заключающих в себе все возможные комбииации гамет, 9 должны быть сераго цвета, 3 – чернаго, 4 – альбиноса. Согласно представленной таблице альбиносы должны отвечать формулам: 1 ААСС, 2 ААСЧ, 1 ААЧЧ. Если это верно, то при скрещивании альбиносов с черными мышами можно предсказать состав потомства. Опыт был произведен и оказался согласным с теориею.
В этом опыте открывается некоторый новый элемент. Оказывается, что один фенотипический признак – черный цвет, серый цвет – можетъ быть результатом совместнаго действия двух факторов, которые, будучи разделены и являясь изолированными, не производят никакого действия.
Сторонники гипотезы „присутствия“ и „отсутствия“ объясняют случай с черными мышами несколько иначе, чем Кено. Они выдвигают три фактора, которые потом сводят к двум. Факторы эти: X – хромоген, противоположно ему х – отсутствие хромогена, С – серый цвет, противоположно ему с – отсутствие сераго цвета, Ч – черный цвет, противоположно ему ч – отсутствие чернаго цвета. Но последнее отсутствие сторонниками излагаемой гипотезы отрицается. Они предполагают, что признак черноты имеется в каждом из родителей, т. е. имеется во всех мышах. Серый цвет является доминантным по отношению к черному. Не смотря на то, что действуют три фактора, скрещивание будет дигибридным, так как один фактор (Ч) неизменно будет присутствовать во всех соединениях и комбинироваться будут только X и С, то разделяясь (Хс, Сх), то соединяясь (ХС), то исчезая совсем (хс). Бетсон серый цвет, как доминантный по отношению к черному, назвал эпистатическим, а черный по отношению к серому гипостатическим. Ясно, что и по этому объяснению второе поколение мышей выразится формулою 9С, 34, 4А.
Из гипотезы, что окраска особи обусловливается совместным действием двух факторов – хромогена и фактора красящаго – следует, что иногда от скрещивания альбиносов может получиться цветное потомство. Альбинизм обусловливается 1) отсутствием хромогена, 2) отсутствием красящаго фактора и 3) отсутствием обоих. Если будет произведено скрещивание 1 (отсутствие хромогена и присутствие красящаго фактора), со 2 (отсутствие красящаго фактора и присутствие хромогена), то в потомстве будет и хромоген и красящий фактор и потомство будет цветным. Для проверки этого предположения производились опыты с душистым горошком. Скрещивались особи с белыми цветами, иногда потомство было исключительно альбиносным, но иногда потомство оказывалось с окрашенными цветами. Понятен первый случай (комбинации 1 и 3, 2 и 3; 3 и 3). С точки зрения изложенной гипотезы понятен и второй (комбинации 1 и 2).
По мере того, как производились опыты, выступали все более сложные факты и явления, которые на первых порах казались противоречащими принципам менделизма, а потом блестящим образом подтверждали их.
Кено производил скрещивание серых мышей с мышами шоколадной окраски. Первое поколение было исключительно серым. Но во втором поколении кроме серых и шоколадных мышей явились не только мыши черные, как это было в аналогичных опытах, но еще мыши серо-золоти-стой окраски или темно-желтой. Формула для этого поколения была такою: 9 серых, 3 черных, з золотистых, 1 шоколаднее.
Кено так объяснил это явление. Он предположил, что в определении окраски действуют три фактора: Хромоген (X), обусловливающий возможность окраски, фактор красящий серый – (С, доминантный), или черный (Ч, рецессивный) и кроме этого фактор Т или фактор 0, первый доминантный, сообщающий густоту (значит темнящий) серой или черной окраске, второй, напротив, рецессивный делающий окраску более светлою (осветляющий). Серая родительская мышь имеет формулу ХСТ, а мыш черная – ХЧТ; шоколадная окраска получалась от совместнаго действия с факторами ХЧ фактора 0, серозолотистый нюанс являлся от совместнаго действия того же фактора 0 с факторами ХС. Если это так, то скрещивание серой мыши с шоколадною дает (ХСТ СЧО) серую мыш. Здесь образуется четыре вида гамет ХСТ, ХЧО, ХСО, ХЧТ, или, если отбросить X, как признак общий, будут гаметы СТ, 40, СО, ЧТ. ІІредставив себе шестнадцать возможных комбинаций этих гамет, получим, что в потомстве должно быть 9С, 34, ЗЖ (золотистых), 1Ш (шоколадная). Этот теоретический вывод согласуется с опытом.
Другие ученые (Дюргем, Кестл) объясняли этот факт гипотезою присутствия – отсутствия факторов. Но у них теоретически получились цифры не соответствующие опыту: 12 серых, 3 черных, 1 шоколадная. Тогда некоторые авторы (Бетсон) прибегли к вспомогательной гипотезе. Они приняли, что С (фактор сераго цвета) дает полный эффект только при соединении с черным, при отсутствии чернаго он дает более светлый нюанс, который и есть серозолотистый. Тогда от 12 серых нужно будет отбросить 3 и получим формулу соответствующую опыту.
Опыт представил факты еще более сложные. Одним из самых простых случаев этого рода можно считать появление во втором поколении того, что Бетсон назвал подчиненными типами.
Скрещивание альбиноснаго дутистаго горошка в некоторых случаях дает первое поколение с пурпуровыми цветами. Тогда во втором поколении получаются растения с пурпуровыми цветами, красными и наконец, с белыми. При чем два первые типа являются – каждый – в трех разновидностях. Чтобы выяснить, в чем заключается их различие, нужно припомнить строение гороховаго цветка. Горох принадлежит к семейству мотыльков (раріlіоnaеаe). Название произошло от формы цветка. Задний лепесток у этих растений сильно превышает остальные по величине. Он называется флагом или парусом. Два боковых лепестка называются крыльями или веслами; остальные два лепестка, которые обыкновенно срастаются своими нижними концами, носят имя лодочки. В бутоне флаг обхватывает оба крыла, а крылья обхватывают лодочку. Разновидности nypnypoSaro и краснаго гороха, являющиеся во втором поколении исследуемаго случая, таковы: 1) флаг окрашен сильнее, чем крылья, 2) крылья окрашены сильнее, чем флаг, 3) весь венчик представляет только едва намеченную пурпуровую или красную окраску. Эти разновидности представляют собою подчиненные типы Бетсона. Они являются в правильных и постоянных числовых пропорциях. Для объяснения этого факта Бетсон предположил кроме факторов X (хромогена), К (дающего красную окраску), П (дающего пурпуровую окраску) еще два новых фактора, один из которых определяет бледную окраску крыльев (отсутствие его делает весь венчик окрашенным однообразно), другой определяет всю окраску цветка (отсутствие его делает весь венчик слабо окрашенным). При таком представлении дела теоретические предположения об окраске гороха совпадают с данными опыта.
В данном случае приходится иметь дело с семью разновидностями. Но имеются факты гораздо более сложные. Множество новых форм во втором поколении получалось от скрещивания mirabilis (опыты Корренза), primula farinosa (опыты Грегори), antirrhinum (опыты Баура, г-жи Уэль-дель). Mirabilis, это – ночная красавица, о которой уже была речь. Primula farinosa, это – первоцвет или белая буквица. Цветок у ней не белый, а лиловый (пятилепестной); названа она так, потому что листья у ней, зеленые сверху, снизу как бы покрыты мукой. Растет на болотах и торфяных почвах, цветет в апреле и мае. Antirrhinum, это – львиный зев наших садов. Растение названо так за форму цветка. На наших полях везде можно видеть разновидность antirrhinum – так называемую льнянку. Желтый цветок ее, как и всех antirrhinum, напоминает рот, а названа она так должно быть за сходство ее стебля и листьев со льном (у цветов нет ни малейшаго сходства).
Баур скрещивал разновидность львинаго зева, имеющаго желтые цветы, с красноцветною пелорическою разновидностью. ІІервая разновидность была нормальною, двугубою. Вторая, называемая пелорическою, представляет исключительное явление, ее венчик заканчивается пятью лепестками снабженными каждый – чипорцею. От скрещнвания в первом поколении явились исключптельно нормальные цветы красной окраски с жилками окрашеныыми более густо. Второе поколение поразило Баура своею причудливостью. Он насчитал 20 различных типов, отличающихся частию окраскою и частию симметриею. Руководясь гипотезою присутствия – отсутствия, он предположил 10 различных факторов, комбинации которых произвели 20 различных типов. Определяющиеся его предположениями числовые пропорции отвечают действительности.
Опыт навел менделистов на противоположные факты. Доселе была речь о том, что один признак оказывался в сущности не одним и производился многими факторами. Теперь должно указать противоположные явления: несколько различных признаков, оказывается, в некоторых случаях производятся одним фактором. Это открывается из того, что эти признаки никогда не расщепляются. Так серая масть у мышей обыкновенно связана с темным цветом глаз, альбинизм обыкновенно связан с красными глазами.У antirrhinum – львинаго зева некоторые виды окраски одновременно являются и на лепестках и на листъях, двубоковая симметрия венчика оказывается принадлежащею и плоду. Но здесь собственно не представляется чего-либо удивительнаго. В данных случаях дело идет об одном общем признаке – пигментации или симметрии, которые зараз появляются в различных органах животнаго или растения. Бывают случаи иного порядка.
Производя многочисленныя скрещивания овса, Нильссон-Эле между прочим изучил две родительские разновидности, которые имели колосок (частичное соцветие, из которых состоит колос) с двумя зернами и отличались одна от другой следующими особенностями, Разновидность а: каждый прицветник имеет ость, ости согнуты, основание колоска имеет волоски, колоски ломки. Разновидность δ: один из прицветников имеет ость, она не согнута, колосок не имеет волосков и не ломок. Первое поколение, получившееся от скрещивания этих разновидностей, представляло собою по отношению к родительским формам посредствующий' тип. Четыре пары аллеломорфных признаков имеются у этих разновидностей. Повидимому имеется на лицо факт полигибридизма и во второмъ поколении должно ждать появления нескольких различных форм. Но три четверти второго поколения оказались типа а, одна четверть – типа δ. Значит, здесь имеется моногибридизм, различные признаки не расщепляются, они оказываются неразрывно связанными между собой. Такое явление, в котором несколько различных признаков производится одним фактором и эти признаки оказываются неразрывными между собою, получило название корреляции.
Нильссон-Эле в своих опытах встретил иные случаи, где два или более факторов производили один признак и, действуя совместно, только усиливали его интенсивность. Он скрещивал разновидность овса, характеризующуюся черною мякиною с разновидностью, имевшею мякину светлую. Первое поколение состояло исключительно из: колосов с черной мякиной. Нужно было ожидать, что три четверти второго поколения будет иметь тип домянантный: а одна четверть – тип рецессивный. Оказалось не то Нильссон-Эле нашел, что на 15 доминантов приходится один экземпляр рецессивный. Он пришел к заключению, что здесь имеется случай дигибридизма. Объяснил он его: так. Он представил, что здесь в одной из родительских сторон действуют два фактора чернаго цвета Ч1 Ч2, другая родительская сторона оогласно гипотезе, присутствия – отсутствия характеризуется отсутствием этих факторов , Гибрид Чі чі Чз чг производит четыре вида гамет: ЧіЧг, Чічг, чіЧа, чіЧ2. Производя всевозможные комбинации этих гамет по два, получим, что из 16 комбинаций в 15 должен быть фактор чернаго цвета, т. е. получим то, что получил Нильссон-Эле.
Нильссонъ-Эле имел дело с еще более сложным случаем, Он скрещивал разновидность овса с красными зернами с разновидностью имевшею белые зерна. Дервое поколение все было типа красного, второе поколение оказалось тоже все типа краснаго. Нильссонъ-Эле стал повторять опыты. Иногда во втором поколении являлись некиторые экземпляры с белыми зернами. При продолжении опытов в третьем поколении тип рецессивный, являлся. Нильссон наконец установил такуй формулу, что во втором поколении на 63 растения с красными зернами приходится одно растение с белыми зернами. Руководясь гипотезой присутствия – отсутствия, он объяснил этот факт так. Три фактора производят красную окраску – К1К2Кз совместным действием усиливая один другого. Им соответствует в другой разновидности отсутствие этих факторов – К1К2К3. Краснозерный овес может иметь своими формулами K1K2K3 или К1К3кз или К1К2Кз или К2к1кз. Различное сочетание факторов объясняет различие в интенсивности окраски, а исследование возможных комбинаций факторов показывает, что из 64 случаев в 63 будет присутствовать фактор К.
Викгор Грегуар суммировал изученные и истолкованные из принципов менделизма факты наследственности.
1) У растений. Менделисты много занимались изучением окраски и формы цветов. Вообще пигментация доминанты по отношению к альбинизму и нормальная форма строения доминанта над формами анормальными (двугубые цветы доминантны над пелорическими, простые цветы над двойными).
Расщепление признаков также было предметом тщательного исследования у листвы (зеленая листва доминантна над листьями пестрыми, листья нормальные доминантны над листьями разрезными, листья глубоко зубчатые доминантны над листьями мало выемчатыми), у ствола (высокий ствол доминантен над карликовым, ветвистый ствол над простым) у поверхности органов (эпидермические волоски доминантны над их отсутствием), у плода и зерна (крахмаловидные и круглые зерна доминантны над сахаристыми и морщинистыми, доминантна форма крахмальных зерен – простых и продолговатых или сложных), у колоса злаков (колос без остей у пшеницы доминантен над колосом с остями). Некоторые физиологические свойства оказались подчиненными законам Менделя. Долгожизненность растений (двухгодичность доминантна над одногодичностью, отсутствие сопротивления к растительной рже – ржа есть болезнь хлеба – доминантна над имунитетностью в этом отношении).
Корренз скрещивал маис, имеющий синие и морщинистые зерна, с маисом, имеющим зерна желтка и гладкие. Колос, происшедши от такого оплодотворения. В первом поколении дал исключительно зерна синие и гладкие. Синий и гладкий значит доминантны по отношению к желтому и морщинистому. Следующее поколение ясно показало дигибридность скрещивавшихся разновидностей. Здесь явились зерна би(бисъ) доминантные – синие и гладкие, бирецессивные – желтые и морщинистые и два сорта смешаных – желтые гладкие и синие морщиннстые. Числовые отношения этих зерен между собою можно предсказать наперед: 9 бидоминантных, 3 и 3 смешанных, 1 бирецессивдый.
2) У животных. Изучение окраски представило много примеров подчинения ее законам Менделя. Пигментация вообще доминантна над альбинизмом. Изучение щерсти и оперения показало, что короткая шерсть доминантна над ангорскою – длинною. Много занимались изучением строения гребня у кур. Установили, что гребень розовидный, имеющий плоскую покрытую сосолками поверхность и направленное назад острие доминирует над простым высоким гребнем, имеющим болышиее зубцы. Другой тип гребня – гороховидный с низким средним и с двумя хорошо развитыми боковыми валиками также доминирует над простым. Производили скрещивание розовидного и гороховидного гребней. Так как здесь имелось две особи с одной парою аллеломорфных признаков, то следовало ожидать, что в первом поколении явятся исключительно доминантные особи, а во втором и доминантные и рецессивные в отношении 3:1. Получилось не то, скрещение роговидного и гороховидного гребня дали в первом поколении новый тип гребня, который назвали ореховидным в виду его сходства с половинкою грецкаго ореха. Во втором поколении явились формы ореховидная (9), розовидная (3), горовидная (3) и еще простая (1). Полученные численные отношения 9, 3, 3, 1 показывают, что здесь имеется налицо случай дигибридизма – гребень создается не одним, а двумя могущими разделяться факторами. Изучение рогов у скота показало, что отсутствие рогов доминантно над ненормальным присутствием рогов. Из физиологических признаков – обыкновенная походка мышей доминантна над танцующею походкою особой породы японских мышей. Из признаков ненормальных полидактилизм – образование многих пальцев – доминантен над нормальным количеством пальцев.
Над животными производилось и производится безчисленное количество опытов. Обширное обозрение их дано в экспериментальной зоологии Моргана. Здесь находим кроме ранее указанных опытов разнообразнейшие опыты с мышами, с морскими свинками, с кроликами, с крысами, с кошками, лошадьми, овцами, кроме опытов с гребкями опыты с лапами, оперением и хохлами кур, опыты с голубями, опыты с пигментом раковин у улитоъ helix hortensis и helix nemoralis, опыты с шелковичными червями, с жуками, с бабочками, с лягушками, с морскими ежами. Во всех этих опытах руководились принципами Менделя и в общем получались результаты согласные с этими принципами.
3) У человека. В вопросах о наследственности производит эксперименты над людьми нельзя, но можно производить наблюдения. Таковые и производились. ІІользовались семейными статистиками. Получать цифры точно соответствующие менделевским пропорциям здесь было невозможно, потому что менделевские цифры получаются при большом количестве случаев, какъ цифры определяемыя теориею вероятностей, а в наблюдениях над людьми приходилось иметь дело с единичными или с немногими случаями. Сравнительно больше изучена теперь многопалость – явление довольно нередкое среди людей. Если полидактилия доминантна, а нормальное строение рецессивно, то представляется вероятным, что многопалый родитель является наследником многопалаго и нормальнаго родителя и что следовательно половые клетки его заключают и М (многопалость) и Н (нормальность). При браке его с нормальным лицом представляется вероятным, что половина детей будет нормальными, многопалыми. В одном случае это так и оказалось. Родилось четверо нормальных и четверо многопалых детей. Бывали случаи, что при одном из родителей многопалом и при другом нормальном все дети рождались многопалыми. Здесь налицо доминантный характер многопалости. Но известны другие случаи, где многопалость оказывается имеющею характер рецессивный, а нормальное количество пальцев характеризуется доминантностью. Так нормальный мужчина женился на особе имевшей шесть пальцев на левой руке. От них родилось восемнадцать человек детей, и только один ребенок оказался ненормальным.
Доминантность пигментации над альбинизмом можетъ быть наблюдаема у негров. Рождаются негры белые – альбиносы. Они не похожи на белых людей кавказского племени, так как их белизна обусловливается не окраскою, а отсутствием пигмента. При браках с нормальными неграми эта белизна оказывается рецессивной.
Наблюдения над глазами показали, что темные и черные глаза доминируют над серыми и голубыми. Далее оказалось, что некоторые аномалии или патологические расположения являются доминантными по отношению к нормальным. Так, руки с короткими пальцами доминируют над руками с нормальными пальцами, катаракт доминирует над нормальным глазом.
Но вообще наблюдения над людьми не могли представить много решающих данных. Наблюдавшиеся явления были сложны, а случаи наблюдений были немногочисленны. Одно только можно установить с несомненностью, что поскольку эти явления поддавались изучению, все заставляет предполагать, что они подчиняются законам Менделя.
Таковы важнейшие данные опытов и наблюдений по методу Менделя над растениями, животными и людьми.
Но опыты показали далее, что в природе существуют и явные уклонения от менделевских формул. Возможны два объяснения этих явлений: или 1) что менделевские формулы не имеют всеобщаго характера и приложимы только к некоторым группам организмов или 2) что в случаях представляющих безспорное несогласие с менделевскими формулами действуют некоторые сиецифические факторы, которые парализуют действие менделевских законов. В природе такие случаи встречаются постоянно. Ни один естественный закон не имеет возможности проявить себя вполне. Ни одна планета не движется согласно законам Кеплера. Гора препятствует земле проявить вполне свою притягивающую силу по отношению к маятнику. Местный магнетизм нарушает правильность в направленииг космаса. Так и в области биологии. Менделисты, понятно, принимают второе предположение. С Бетсоном во главе они разделили факты, не подчиняющиеся законам Менделя, на две категории. В одной категории они предположили по английской терминологии „gametic coupling“ а в другой „ga-metic repulsion“. Ha русский язык эти термины переводят „сочетание гамет“ и „отталкивание гамет“ или, что тоже, „сочетание факторов“ и „отталкивание факторов“. Перевод, конечно, буквальный. Но согласно смыслу гипотезы вместо слово „сочетание“ понятнее будет слово „притяжение“ (attraction). Дело идет о том, что между некоторыми факторами определяющими наследственност обнаруживается взаимное притяжение, стремление к совместности в действии, между другими факторами открывается обратное явление – стремление к разъедннению, к обособленной друг от друга деятельности.
Бетеон производил опыты над Latlivrus – разновидностямя дупшитаго горошка (у нас Lathyrns растет около заборов, на лугах, на полях особенно, где ячмень). Он скрещивал две белоцветныя разновидности. Первоё поколение оказалось однообразно составленным из растений с пурпуровыми цветами. Во втором поколении на 27 растений с пурпуровыми цветами пришлось 9 – с цветами красными, 28 – с белыми, т. е. на 36 цветных растений пришлось 28 белых или иначе на 9 цветных – 7 белых: но это явно отвечает формуле Менделя 9, 3, з, 1. Но формулам Менделя оказалось несоответствующим соотношение между окраскою и зернами пыльцы. Скрещивавшиеся альбиносные расы различались между собою формою зерен пыльцы: у одной расы зерна были круглые и плоские вроде лепешекф, а у другой – продолговатыя, цилиндрообразныя.
Первое поколение (пурпуровое) имело исключительно пыльцу продолговатой формы – длинную. Значит длина есть доминирующий признак. Во втором поколении согласно теории Менделя в цветах каждаго типа окраски должно бы было быть три четверти длинных и одна четверть круглых пыльцевых зерен. У растений альбиносных это и оказалось. На ЗД приходилось 1К (з – длинных, 1 – круглое). Но у пурпуровых цветов на 12ПД пришлось одно ПК, у красных цветов на 3, 2 Кр К (буквами Кр я обозначаю красные, буквою К – круглые) пришлось 1 Кр Д. Здесь возобладал рецессивиый признак. Действительные данные опыта были таковы. 1528 ДН (буквою Н я обозначаю не красные цветы) 106 КН, 117 Кр Д, 381 Кр К.
В общем второе поколение оказываетоя приблизительно состоящим из трех четвертей растений с длинною пыльцею и одной четверти с круглою. Но оказывается, здесь нельзя допустить независимаго расщепления соединенных вместе факторов. В выше излагавшихся опытах факторы в различных комбинациях расходились и сходились совершенно свободно. Здесь нет этого четыре возможные комбинации НД, НК, КрД( КріК должны бали бы быть но количеству равными между собою. Но на самоме деле комбинации НД и КрК преобладают над другими. Бетсон принял, что существует некоторое притяжение между факторами Н и Д, происходит сочетание этих факторов и оно влечет за собою преимущественное образование гамет НД и соответственно образование гамет КрК. Преимущественное, но неисключительное. Имеются растения КрД и Η К.
Бетсон пошел дальше. Он показал, что сочетание факторов, о которых идет речь, соответствует определенным числам. Если принять, что взаимно притягивающиеся гаметы сочетаются в 7 раз чаще, чем гаметы не обладающие свойством притяжения, то для гаметы получится следующая формула: 7НД, 1HK, 1КрД, 7КрК, Все эти комбинации осуществленные в 256 растениях приводят к формуле 177НД, 15HK, 15КрД, 49КрК. Менделисты сближают эту формулу с полученными из опыта цифрамн 1528HK, 106KH, 117ХрД, 318КрК.
Бетсон производил еще скрещивание разновидностей Lathyrus, из которых у одной пыльники характеризовались плодородием, у другой – скудостью, загибы листьев у первой имели густую окраску, у второй были светлыми. Обозначим плодородные пыльники буквою П, скудные – буквою М (малоплодные), густую окраску буквою Г, светлую – С. Состав первого поколения тогда охарактеризуется буквами ПГ. Плодородие и густота доминантны над скудостью и светлой окраской. Второе поколение отвечало формуле 627 ПГ, 27МГ, 17ПС, 214MC. Преобладание ПГ над і\1С заставляет предположить притяжение между факторами П и Г. Но состав поколения может быть объяснен только новою пропорциею гамет: 15ΠΓ, іМГ, іПС, 15MC. Вычисление возможных комбинаций между такими гаметами дает такую формулу для второго поколения: 637ПГ, 27МГ, 27ПС, 194MC. Формула эта близка к тому, что дает опыт.
Скрещивание двух разновидностей гороха, различающихся между собою. присутствием (П) или отсутствием (0) усиков и круглою (К) формою зерен в одной разновидности и угловатою (У) в другой, дали во втором поколении растение отвечавшие такой формуле: 319ІШ, 40К, ЗПУ, 1230У. Это приводит к новой формуле сочетания различных гамет. Если между ними допустить такое соотношение: 63ÜK, ІОК. ІПУ, 630У, то из числа экземпляров соответствующаго имевшему место в опыте должно получиться: 833ПК, 3, 40К, 3, 4ПУ, Ю90У. Эти цифры приближаются к действительной пропорции.
Скрещивание пурпуроцветнаго Lathyrus, имеющаго у цветка поднятый парус (Р, развернутый), с белодветною разновидностею (Б), имеющаго парус свернутый (С), для второго поколения дало такой состав, который требует следующаго распределения гамет: 127ПР, іПС, іБР, 127БС.
Обратили внимание на своеобразные свойства чисел, которыми при различных скрещиваниях у различных растений определяется взаимоотношение гамет. 127=(2х63)+1; 63=(2х31) + 1; 31=(2х15)+1; 15=(2х7)+1; 7=(2хЗ)+1; 3= (2х1)+1. Член левой части каждаго из этих уравненей есть изменяющийся множитель второй части предыдущаго уравнения и он всегда равен левому члену предыдущаго уравнения уменьшенному на единицу и разделенному на 2.
Имея в виду эти цифры, Бетсон предположил, что полная серия гаметических. формул должна быть такою.
1, 1,1, 1 (нормальный случай; притяжения факторов нет).
3, 1, 1, 3 (Бетсоном еще не наблюдавшийся случай).
7, 1, 1, 7 (наблюдавшийся случай).
15, 1, 1, 15 (наблюдавшийся).
31, 1, 1, 1, 31 (еще не наблюдавшийся).
63, 1, 1, 63 (наблюдавшийся).
127, 1, 1, 127 (наблюдавшийся).
Грегори для китайскаго первоцвета (близкаго к primula farinosa – белой буквице) нашел формулу 7, 1, 1, 7. Ваур при скрещиваниях antirrhinum – львинаго зева получил теоретически предсказанную Бетсоном пропорцию 3, 1, 1, 3. Много раз в своих опытах он приходил к формуле 7, 1, 1, 7. Но кроме того он столкнулся с фактами, которые не обнимаются цифрами Бетсона. Общий вид формул Ветсона таков: n, 1, 1, n. Но Баур увидел необходимость для некоторых случаев дать формулу: n, 1, 1, х, где х>n, и опыты Баура иногда приводили к формуле: 7. 1, 1, 9. Кроме того для соотава второго поколения в некоторых случаях он нашел формулу: 4, 1, 1, 4 более подходящею, чем 3, 1, 1, 3.
Сочетание факторов или притяжение само собою предполагает существование отталкивания. Где имеются симиатии, там всегда существуют и антицатии. И из последняго иачала менделисты объясняют многие факты наследственности.
Некоторые разновидности Lathyrus, как уже было говорено, имеют парус совершенно раскрытый и прямой. Есть разновидности имеющие парус согнутый вверху на две стороны в форме капюшона. Бетсон скрещивал прямую белоцветную разновидность с загнутою белоцветною разновидностью. Первое поколение дало пурпуровые прямые цветы. Второе поколение с точки зрения окраски представило три группы (оставляя в стороне типы подчиненные): с пурпуровыми цветами, с красными цветами, с белыми цветами. Из пурпуровых цветов у одних – парус прямой, у других – загнутый. Тоже самое у цветов белых, но все красные цветы имеют парус прямой. Красный цвет не соединяется с загнутым парусом. Как объяснить это? Бетсон предположил здесь действие четырех факторов. Фактор X (хромоген) не дает окраски, но обусловливает ея возможность, ему противоположен 0 (отсутствие хромогена), устраняющий возможность окраски; фактор Кр, совместно с X определяющий красный цвет окраски, ему противоположен Н (нет окраски, даже и в присутствии хромогена); фактор П преобразующий красную окраску произведенную ХКр в пурпуровую, ему противоположен Б (без пурпура); наконец фактор Р (развернутый), ему противоположен С (свернутый). Отсутствие соединения краснаго цвета с согнутостью паруса объясняется тем, что никакая зигота происшедшая от гибридов перваго поколения не самозиготна зараз по отношению к Б и С. т. е. что никакая зигота образовавшаяся при оплодотворении из двух гамет не может иметь формулаши ХХКрКрББСС или ХОКрКрББСС или ХХКНСС. Если бы какие-либо растения удовлетворяли какой-либо из этих формул, они дали бы цветы красные и с загнутым парусом.
Для объяснения того факта, что во втором поколении не оказалось особей удовлетворяющих формул БС БС можно продположить, что были и мужские гаметы Б С и женские, но что они не встретились при оплодотворении. Но это предположение является безусловно невероятным. Все другие комбинации осуществились многократно, а эта не осуществилась ни разу. Невидно никаких препятствий, которые мешали бы встретиться Б с Б или С с С. В красных дветах и в белых имеется соединение Б с Б, в согнутых цветах имеется соединение С с С. Ничто видно не препятствует и встрече Б с С, потому что изучение третьяго поколения показало Бетсону, что все пурпуровые и прямые цветы второго поколения гетерозиготну по отношению к II и Р и имеюг формулою ПБРС. Остается допустить, что гибриды перваго поколения не производят гамет БС и что всякая гамета содержая С долша также имет и П. Числовые величины различных групп второго поколения показывают, что в общем, расщепление каждаго признака из аллеломорфной пары совершается по нормальному образцу. Нельзя допустить, чтобы один класс гамет соверщенно отсутствовал, не предположив в то же время совершеннаго отсутствия противоположнаго ему класса. Если нет БС, то нет ни одной гаметы и IIP. Бетсон и говорит, что между факторами П и Р обнаруживается вид отталкивания, которое и препятствует им встретиться в одной гамете. Эта гипотеза отсутствия гамет ПР и БС не только объясняет, почему во втором поколении все красные цветы имеют прямой парус, но она даетъ возможность дать отчет о детальном составе второго поколения. При четырех факторах X и 0, Кр и Η, П и Б, Р и С и при расщеплении признаков по нормальному типу гибриды имели бы 16 видов гамет. Вычисление показывает, что они должны бы были дать 256 генотипических комбинаций (162), которые распределились бы в 6 фенотипических формах: 1) пурпуровые прямые, 2) пурпуровые загнутые, 3) красные прямые, 4) красные загнутые, 5) белые прямые, 6) белые загнутые. Но если ПС или БС ничего не производят, то остается только 8 видов гамет, которые дают 64 генотипические комбинации (88), распределенные между пятью фенотипическими (из перечисленных выше шести групп нужно исключить четвертую: красные загнутые). Теория 64 генотипических комбиыаций и 5 фенотипических групп дает числа весьма близкие к тем, которые Бетсон получил в опыте.
Изучая третье поколение, происшедшее от пяти фенотипических групп второго, Бетсоп получил результаты, которые определялись заранее его гипотезою генотипическаго состава гамет второго поколения. Все растения с пурпуровыми и прямыми цветами дали потомство с диссоциированными П и Р. Все они характеризовались формулою ПБРС. Это и должно быть, если гаметы ПР не соединяются.
Подобные явления отталкивания наблюдались и у некоторых животных. Из растений Грегори наблюдал их у первоцвета (буквицы) и Баур – у львинаго зева и водосборов (в некоторых местах у нас называемых колокольчиками).
Удалось установить в некоторых случаях, когда действует притяжение и когда отталкивание факторов. Выше была речь о скрещивании Lathyrus с плодородными пыльниками и густой окраской (П и Г) с Lathyrus с малоплодными пыльниками и светлой окраской (М и С). Бетсон произвел скрещивание между ПС и МГ. Если бы явления протекали согласно формулам Менделя, то старое поколение происшедшее от этих гибридов дало бы четыре фенотипические формы: ПГ, ПС, МГ, MC. В действительности последней формы не оказалось совсем. Для объяснения явления Бетсон предположил отталкивание между факторами – плодородный и густой (Π. Г.). При скрещивании ГІГ и MC факторы П и Г подчинялись закону взаимнаго притяжения, но при скрещивании ПС и МГ между ними проявилось отталкивание. В первом случае оба фактора II Г были соединены в одном из двух родителей, произведших гибрида, во втором случае эти факторы разделены в отцовском и материнском организмах. Бетсон вывел следующее правило: если A, а и В, b являются двумя парами аллеломорфных признаков подчиненных закону притяжения и отталкивания, то факторы А и В будут взаимно отталкиваться при соединении гамет типа Ab и аВ и будут взаимно притягиваться при соединении гамет типа AB ii ab. Иначе это правило формулируют так: в некоторых случаях полигибридизма гаметы в очень большом числе (при притяжении) или даже исключительно (при отталкивании) дают рождение гаметам аналогичным с теми, которые им дали рождение. Это правило подтверждалось у Бетсона во всех его опытах. Баур с своей стороны дал ему новые подтверждения.
Бетсон и другие менделисты для объяснения только что описанных фактов предполагают существование двух факторов – притяжения и отталкивания. Но Грегуар допускает, что все эти явления могут быть объяснены из одного притяжения при допущении гипотезы „присутствия – отсутствия“. Он обращается к явлениям у Lathyrus с плодоносными пыльниками и густо окрашенными загибами листьев и Lathyrus с малоплодными пылъниками и светло-окрашенными загибами. Если родители полигибридов имеют формулами ПГ и MC, тο существует притяжение между П и Г с одной стороны и М и С с другой, но притяжение это действует не с абсолютною силою, вследствие чего является небольшое число гамет ПС и МГ. Если родители полигибридов будут иметь формулами ПС и МГ, то получится потомство совершенное не имеющее экземпляров отвечающих формуле MC. Объяснить это можно 1) или предположением, что между П и С, равно как и между М и Г существует абсолютное и исключительное притяжение, препятствующее образованию гамет типа MC:
2) или предположением, что в полной серии гамет: η ПС. іПГ, ШС, η МГ, η так велико, что практически оно приводит к совершенному отсутствию гамет ПГ и MC. Так те явления, которые Бетсон объясняет отталкиванием, Грегуар хочет истолковать, как крайние случаи притяжения. Опытъ уже дал такие пропорции, как 127, 1, 1, 127, естественно предположит, что явятся цифры и большие, чем 127, между тем с точки зрения теории вероятностей, если мы имеем с одной стороны 127 шансов, а с другой 1 (127 белых шаров, один – черный), вероятност одного шанса (вынуть черный шар) приближается к нулю.
Особую группу фактов, которую в настоящее время ученые рассматривают с точки зрения принципов менделизма, представляют явления пола. Общим правилом относительно пола, которое доселе совершенно не имело объяснения, является то, что особи мужского и женского пола рождаются приблизительно в равном числе. Наблюдения установили еще, что иногда замечается передача отцовских особенностей женскому потомству, а женских – мужскому.
Опыты над бабочками (крыжовной пядиницей) и над курами (шелковистыми и леггорнами) установили еще, что, свойства потомства обуславливаются полом разновидностей (явление совершенно противоположное тому, что получил Мендель в Pisum sativum): от скрещивания самцев крыжовной пяденицы окрашенных более ярко (abraxas grossu-lariafa) с самками бледной окраски (abraxas lacticolor) происходит не такое потомство, которое получается от скрещивания бледных самок с яркими самцами.
Равенство полов представляет собою такое явление целесообразности, которое нередко вводили в состав телеологическаго доказательства бытия Божия. Противники телеологии всегда возмущались этим. Не умея найти естественного объяснения для факта, они настаивали на том, что он имеет естественную причину. Противники телеологии не имеют понятия о том, что такое телеология. Ни один телеолог не представляет, что Бог непосредственною силою уравнивает возмущения планет, уравнивает число особей мужского и женского пола, создает непосредственно организмы приспособленные к среде или Сам непосредственно приспособляет среду к организмам. Нет теологи признают, что все эти явления имеют для себя естественные причины, но что целесообразная совокупность этих естественных причин имеет свою последнюю причину в Боге. Непосредственное воздействие Божеетва на мировой строй имеет место там, где этот строй расстраивается противоестественным фактором – злою волей свободных существ, или там, где добрая воля таких существ ищет опоры и содействия в своем стремлении к совершенствованию.
Численное равенство особей мужскаго и женскаго пола должно иметь для себя естественную причину. Менделевская теория гамет показывает возможность существования такой причины и подсказывает сущность объяснения. Формы предложенных на менделевской основе гипотез различны, но сущность их одна. Простейшая из них принадлежит Бетсону. Обозначим, чрез М большое мужской фактор, чрез m малое – отсутствие мужскаго фактора, через F большое – женский фактор, через f малое – отсутствие женского фактора. Бетсон предполагает, что строение самца отвечает формуле Mmff, строение самки – Ffmm. При соединении самца с самкою образуются такие сочетания гамет:
| Гаметы от самца | Гаметы от самки |
| Mf | fm |
| mf | Fm |
| Mf | Fm |
| mf | fm |
Понятно, что последняя комбинация гамет должна дать безплодные резулътаты, но такие же результаты дает предпоследняя комбинация. М и F, встретившись в одной зиготе, парализуют друг друга взаимно. Зиготы Mf Fm и mmff не могут развиваться. Новые особи образуются из зиготы Mmff и Ffmm, при чем теория вероятностей устанавливает что при большом количестве соединений те и другие зиготы образуются в равном числе, и следовательно число самцов и самок должны быть равны между собою30.
В природе замечается тенденция передачи некоторых отцовских свойств женскому поколению и материнских мужскому. Под этими свойствами нельзя разуметь тех, которые фактически связаны с полом, хотя сами и не имеют полового характера. Бородатость отца не передается дочерям. В породах безрогих Овец, рогатые бараны не производят овец рогатых. Но есть иные свойства, которые безразлично могут принадлежать и тому и другому полу. Я лично знаю две семьи, в которых глухота отцов в значительной степеди передалась дочерям, но не отразилась заметным образом на сыновьях. Здесь, исходя из принципов Менделя, можно объяснять явление так. Признак глухоты, передаваемый от отца вероятно в какой-нибудь неразъединимой комбинации, может быть несовместим или по крайней мере трудно совместим с другими признаками, присущими сыновней зиготе, но он совместим с свойствами дочерней зиготы. Отсюда у некоторых дочерей (не у всех) этот признак проявится.
Факт, что отцовские свойства порою передаются дочерям, а дочерние – сыновьям уже определяет, что пол сказывается на свойствах потомства. Влияние пола скрещиваемых разновидностей было предметом экспериментальных изследований. Так густо окрашеннаго самца крыжовной пяденицы – grossulariata – скрещивали с бледноокрашенною самкою – lacticolor. Первое поколение все было grossulariata. Второе поколение дало то, что следовало ожидать по формуле Менделя: на 3 grossulariata пришлось 1 lacticolor. При чемъ все lacticolor были самки. Это уже показывало на то, что явление сложнее, чем те, которые определяются элементарнейшей из формул Менделя. Было произведено скрещивание самки lacticolor с самцем grossulariata, происшедшим от первого поколения. Явились и самцы и самки lacticolor и самцы и самки grossulariata, при том все четыре группы в равном количестве. Шлученнаго таким образом самца lacticolor скрестили с самкою перваго поколения. Lacticolor и grossulariata явились в равном количестве, но все нормальные grossulariata были самцами, все lacticolor были самками. Этот факт объясняют таким образом. Признак g (grossulariata) доминирует над 1 (iacticolor). Доминирующий фактор у самки F (женский) гетерозиготен, тоесть у ней имеется и этот фактор и то, что называется отсутствием фактора, – Ff. У самца имеем ff. Между Gr и F в зиготе гетерозиготной в отношении обоих этих факторов, т. е. имеющей и фактора и его отрицание существует отталкивание. Такие зиготы (Ff, Gg) всегда должны быть женского пола и они должны давать равное число гамет Fg и fG.
Еще были произведены опыты скрещивания шелковистой породы кур, характеризующихся чрезвычайным обилием пигмента, с коричневыми леггорнами, у которых этот пигмент отсутствует. Скрещивание петуха леггорна с шелковистой курицей дало в первом поколении безпигментное потомство. Происшедшее от перваго поколения второе дало особи с пигментацией, без пигмента и промежуточные формы с различными степенями пигментации. При чем были куры с сильной пигментацией, но петухов с вполне нормальной пигментацией не было. Было произведено обратное скрещивание – шелковистого петуха с курицей леггорном. Петухи первого поколения оказались безпигментными, но куры были почти нормально пигментированы. Свойство отца передалось дочерям. Этот случай уже былъ истолкован. Последующее поколение заключало в себе и самцев и самок пигментированных, без пигмента, с различными степенями пигментации. Этого и следовало ожидать.
Явления наследственности у шелковистых кур и у леггорнов навели Пеннетта на очень ядовитую мысль. Он отмечает, что фактор женскаго пола иногда оказывается несовместимым с двойною дозою какого – либо другого фактора, и говорит: „возможно, что некоторые из свойств, которыми мужчина отличается от женщины, покоятся на такого рода различии. Некоторые интеллектуальные качества, например, могут зависеть от существования в особи двойной дозы фактора, отталкиваемого фактором женского пола. Если это так, и если женщина стремится к достижению цели, требующей таких интеллектуальных качеств, то ей не помогут ни воспитание, ни тренировка. Задачей ее будет ввести фактор, от котораго зависят эти качества, в яйцо несущее одновременно и фактор женскаго пола31“ т. е. задача невозможна. Смысл всей зтой речи конечно тот, что двойная порция ума, которою владеет мужчина, несовместима с женским полом. „Гений и злодейство – две вещи несовместимые“, говорит Моцарт у Пушкина (Моцарт и Сальери), Если это так, то и это будут объяснять из принципов менделизма, а пока, руководясь этими принципами, Пеннетт подсказывает:
Ум и женщина – две вещи несовместные.
Здесь Панетт идет, кажется, гораздо дальше, чем хотел идти Мендель.
Мендель создал свою теорию, когда внимание биологов всего мира было всецело обращено на учение Дарвина. Теперь учение Дарвина исследуется и проверяется не менее внимательно, чем и тогда, но оно давно уже перестало быть окончательным научным откровением. – Много в этом учении оказалось и сомнительного и несомненно неверного. Теперь менделизм наносит этой доктрине жестокие удары. Дарвиновская теория происхождения видов предполагала: 1) непрерывность изменчивости. Ею отрицались скачки в природе. Хобот слона произошел от постепенного удлинения носа его предков, и это удлинение должно было совершаться с такою медленностью, что в смежных поколениях оно было неуловимо. 2) Теория Дарвина предполагала, что веякое изменение в индивидууме может передаваться потомству. Менделизм отверг оба эти положения. Он выяснил, что нужно различать между тем, что особь представляет собою совне и что она таит в себе внутри, нужно различать фенотипические и генотипические свойства. Высокорослый горох может хранить в себе потенцию низкорослого потомства. Мендель и меиделисты представили безчисленное количество опытов, в которых изменения происходили не непрерывно, а скачками, внезапно. При чем эти изменения обычно вовсе не зависели от того, каким превратностям подвергался индивидуум втечение своей жизни. Можно привести пример, характеризующий взаимное отношение дарвиновского и менделевского объяснений. Есть в Африке бабочка amauris (из семейства Danaidae). Повидимому ее вкус отвратителен для птиц и обезьян. Они ее и не трогают. Есть там же бабочка Euralia (из семейства Nymphalidae), она повидимому не имеет неприятного вкуса, но она так похожа по своей окраске на amaurife, что, смешивая ее с amauris, обезьяны птицы ее не трогают. Euralia имеет то, что в зоологии называется покровительственной окраской – мимикрией. По теории Дарвина эта окраска могла быть приобретена лишь втечение громаднаго периода времени путем суммирования мельчайших изменений и на первых порах не могла быть покровительственною, т. е. охраняемою естественным отбором. Ясно, что дарвинизм не может объяснить факта. Но исходя из теории Менделя и производя опыты согласно его принципам, установили, что здесь все совершалось по менделевским формулам, и разновидности являлись так как они всегда являются в менделизме, сразу, внезапно.
Может быть сразу могуг возникать новые виды, роды и типы? Природа втечение многих поколений может вырабатывать фактор, не проявляющий себя вовне, но потом внезапно в известной комбинации гамет становящиеся доминантным и производящий новый вид. Уже исследования Келликера, о которых была речь во второй главе, наводят мысль на эту возможность. Эту возможность подтверждают работы Коржинскаго и Бетсона. Гюго де Фриз одновременно с Коррензом и Чермаком открывший в 1900 году мемуары Менделя, в этом же году и независимо от Менделя дал теорию внезапного возникновения новых видов, теперь тесно сплетшуюся с учением менделизма.
Внезапному превращению одного вида в другой теперь усвоено имя мутации, и теория таких превращений названа мутационной. Сущность ее по де-Фризу такова: органические виды не подлежат непрерывной изменчивости, они втечение более или менее долгих периодов остаются неизменными. Но затем органический вид вступает в мутационный период, организмы прямо производят от себя новые виды. По мнению де-Фриза особенности вида обусловливаются появлением в организме определенных, единичных, резко отграниченных один от другого элементов. Сами эти элементы подлежат изменениям, вариациям, флюктациям, обусловливаемым согласно опытам де-Фриза главным образом питанием, от этого зависят вариации или флюктуации в растениях. Но возникновение видов зависит не от вариаций, а от мутаций. Наступает момент в жизни вида, когда он должен дать жизнь новым видам. По Фризу такой мутационный период пережили в недалеком прошлом многие растения. Такова крупка весенняя, иначе называемая быльник весенний, сухоребрица (Draba тегпа) – маленькое однолетнее растение, принадлежащее к семейству крестоцветных (имеют четыре лепестка расположенных крестообразно), с белыми цветами, лепестки разделяются вверху, плодъ стручек, растет на песчаных местах (6 тычинок, из них 2 больше остальных). У крупки в настоящее время насчитывают до двухсот подвидов, очень сходных, но совершенно самостоятельных, которые поэтому могут быть рассматриваемы как виды. К растениям, подобно крупке недавно пережившим мутационный период, относят подорожники, ивы, шиповник, ежевику. В настоящее время по Фризу переживает мутационный период oenothera Іашаг-kiana (ослинник, онагрик, ночная светильня, двухлетнее растение, семейство onagraricae или oenotherae). Около ста лет назад оно вывезено из Северной Америки и теперь распространилось по Европе. Есть виды oenothera употребляемые в пищу, таков например, корень oenothera biennis, (корень такой же как у моркови) разводимаго в декоративных целях в садах радн его желтых, четырех-лепестковых красивых и сильно пахнущих цветов. Фриз наблюдал oenothera в окрестностях Амстердама, он заметил у этого растения появление новых видов, перенес его в эксперимеытальный сад и втечение многих лет производил над ним наблюдения. Фриз сеял тысячами семена oenothera. Втечение 1889 – 1899 г.г. из этих семян вышло семь новых видов: oenothera gigas, albicla, lata, nanella, rubrinervis. oblonga и scillitans. Последний вид наименее устойчив, только часть его семян дает новый вид. Важно здесь обратить внимание на то, что новый вид возникает не от одного какого-либо организма (oenothera имеют цветы двуполые), а от многих. Семена других растений дают тождественный новый вид. Отсюда следует, что oenothera в потенции заключает в себе возможность мутации в строго определенных направлениях32.
Фриз пришел к своей теории путем исследований над растениями. Теперь мутации находят и у животных. Возможно приложение этой теории, неотделяемой от теории Менделя, и к человеку. Ею устраняется одно из возражений направляемых против библейской истории. В библии возникновение рас, народностей представляется совершающимся как то незаметно и в то же время для этого возникновения Библия повидимому назначает немного годов. С точки зрения доменделевской и дофризовской биологии образование рас так резко расходящихся между собою, как кавказская и негрская, могло совершиться только втечение тысячелетий, Эти тысячелетия мудрено втиснут в рамки библейской хронологии. С точки зрения Менделя и Фриза различные человеческие расы могли возникнуть внезапно и установиться сразу. Факт существования резко различающихся между собою рас оказывается нисколько неколеблющим хронологии, обычно извлекаемой из Библии.
4. Заключение
В менцелизме открывается двойственность. С одной стороны он является учением строго эмпирическим, в сущности точным описанием фактов, из которых с необходимостью следуют новые факты. С другой стороны может показаться, что учение менделизма о факторах наследственности – доминантах и рецессивах, факторах не действующих, но лишь обусловливающих возможность действия других факторов, как, например, хроматин, обусловливающий возможность окраски, о факторах взаимно притягивающихся, о факторах несовместимых, что-такое учение всецело метафизично и даже мистично. Действительно, если пытаться определять свойства этих предполагаемых факторов, решать вопросы о них – представляют ли они собою вещественную субстанцию или энергию, и что это за субстанция или энергия, то менделизм, оставаясь естественонаучным учением, станет в то же вреня и метафизической доктриной. Правда, давно пора перестать смущаться словом „метафизика“ и давно пора догадаться, что все люди метафизики. Но дело в том, что метафизика, как и физика, бывает и хорошею и плохою, и если бы менделисты пытались давать учение о сущности факторов, управляющих наследственностию, они вступили бы на скользкий путь увлечения плохою метафизикою. Но Мендель не вступал на этот путь. Не увлекает он и менделистов. Факторы менделизма R, Ri. R2, это – условные обозначения начал, производящих явления. Генрих Герц (1857 – 1898), расширивший ваши познания об электрических волнах, мечтал о физике без гипотез. Эта физика не должна была толковать о том, что такое свет – особая материя, дрожание эфира или что иное, она должна была бы только математически определять законы действия явлений. Герц невольно всвоминался мне при изучении работ Менделя. В физике мы имеем, что иногда от прибавления света к свету получается мрак – в явлениях интерференции, когда возвышения одних световых волн совпадут с углублениями других. У Менделя мы имеем, что соединение высокорослых родителей иногда может дать низкорослое потомство. Толковать о сущности факторов, производящих эти явления безполезно. Но изучение фактов и их численного взаимоотяошения дает нам формулы теории соединений. Правда, найденные числа не являются коэффициентами бинома Ньютона – в одном случае они представляют собою и эти коэффициенты, только не так поставленные: 3, 1. і, 3 – но ясно, что сущность дела здесь та же. Нам даны комбинации, мы можем проследить, как они образуются, и предвами возникает чисто алгебраическая задача – определять законы образования этих комбинаций и число элементов, из которых они образуются. Эти элементы мы будем обозначать не числами 1, 2, 3, 4 и т. д., а наименованиями – отсутствие красного цвета, присутствие красного цвета, фактор усиливающий окраску, фактор ослабляющий окраску, фактор к двойной дозе, названия иное, но сущность дела та же.
Как в числах раз даны элементы и закон их сочетания, может быть определен результат, так и в мёнделизме результат предсказывается. Менделизм есть прежде всего описание опытов. Далее, менделизм есть переложение словесного описания на язык цифр. Затем оказывается, что раз дано известное количество цифр, то дальнейшие цифры будут представлять собою функцию данных. Опыт ненужен более. Вычисление предскажет нам в цифрах будущие явления, а в конце концов язык цифр мы можем снова переложитъ на язык слов.
Если нельзя узнать сущности фактора, то за всем тем конечно нужно по возможности стремиться исследовать его проявления. Современная биология и не останавливается на внешней стороне явлений менделизма. Цитологическое и эмбриологическое исследование привлекаются для выяснения фактов наследственности. Изучение клетки получило широкое развитие. Много сделано и в областн изучения образования из клетки новых клеток, т. е. произведения клеткою или клетками нового потомства. Исследования под микроскопом показывают, что образование и развитие зародыша совершаются согласно принципам Менделя, наблюдаются даже реальные единицы аналогичные факторам менделизма – зернышки называемые хромозомами33.
Теория Менделя уже имеет и без сомнения будетъ иметь громадное биологическое значение. Как сама в себе, так и в своем отношении к биологии она является как бы двойственной. С одной стороны она предлагает биологам сузить проблему происхождения новых форм. Во второй половине XIX века вопрос о происхождени был поставлен очень широко – решали проблему происхождения всего органическаго мира – растительного, животного, человеческого вместе с такими сложными проявлениями человеческого духа, как наука, мораль, религия, вся культура с ее материальной и духовной стороны. Все поняли Дарвин объяснил миру. Объяснение было до наивности просто. Но что же может быть лучше простых объяснений? Дарвин, говорят, не любил алгебры. На самом деле гораздо проще довольствоваться четырьмя действиями арифметики. Теория Менделя призывает биологов к скромности. Вопрос о происхождении новых форм, о факторах обусловливающих их возникновение, оказывается настолько сложным, что в сущности с научной точки зрения при настоящем состоянии знаний недопустимы широкие обобщения по вопросу о происхождении родов, классов и типов. Решения этих вопросов предложенныя во второй половине XIX века на самом деле столько же научны, как вавилонские или греческие сказания о происхождении растений, животных и людей. Родословные животных и человека, составленныя досужими биологами вроде Геккеля оказались имеющими право претендовать лишь на такую степень достоверности, на какую могут претендовать родословные гомеровских героев. Загробный голос Менделя призвал биологов опьяненных теориею Дарвина к трезвому исследованию явлений. От широкой постановки вопросов нужно отказаться. Разумеется нельзя и не следует запрещать появление в печати трактатов, имеющих заглавие „история органического мира“, но следует вместе с тем позаботиться о таком естественнонаучном образовании общества, чтобы оно понимало, что такие произведения, как истории органическаго мира, относятся более к области изящной словесности, чем строгой науки.
Но если с одной стороны менделизм суживает задачи биологического исследования, то с другой стороны он углубляет их. Точно и строго описывая то, что есть и отказываясь от фантастических описаний того, что было менделизм в то же время стремится истолковать законы того, что есть, т. е. существующаго, так, чтобы вместо прежних фантастических и неопределенных предсказаний будущаго, во многих случаях имет возможность предсказывать это будущее математически точно, так предсказываются теперь лунные и солнечные затмения.
Менделизм начался с того, что монах Мендель в пятидесятых годах прошлого столетия стал производить опыты над горохом. Характер опытов, метод работъ Менделя, намеченные им вопросы, все повидимому указывало для его дела узкие и тесные рамки. Но в действительности оказалось не то. От гороха перешли к наблюдениям и опытам над другими растениями, от растений перешли к животным, от животных перешли и к наблюдениям над людьми, от вопросов о происхождении их физических особенностей перешли к вопросам об их нравственных свойствах. Дарвин от наблюдения над породами голубей перешел к решению проблемы о происхождении религии и морали. Менделизм доселе не ставит дело так широко, да и при тех знаниях, которыми располагает человечество, ставит его так широко нельзя. Но по естественному порядку менделизм не мог ограничиться вопросами о физической наследственносги он должен был заняться проблемами и нравственной наследственности. Между законами наследственности биологической и наследственности моральной нет непереходимой бездны. Если бы между биологическими и нравственными законами наследственности существовали бы только аналогии, и тогда бы менделизм мог оказывать услуги этики. Со времен Эмпедокла сближают любовь с притяжением и отталкивание с враждою. Последнее сближение мне представляется неудачным: вражда и ненависть не аналогпчны отталкиванию; отталкиванию аналогично отвращение. Во всяком случае можно сказать, что между симпатиею и антипатиею с одной стороны и между неразрывностью и несовместимостью факторов (не в абсолютном смысле) существует аналогия. Но у нас дело не в одних аналогиях. Кроме того, что физическою и духовною природою в некоторых отношениях управляют аналогичные законы, между материею и духом существует крепкая связь, и все, что происходит в сфере физической, отражается на сфере духовной и наоборот. Материалист найдет объяснение этого факта в том, что дух есть функция тела. Крайний спиритуалист может выдвинуть тезис, что тело есть функция духа, что дух создает тело сообразно с своими влечениями и стремлениями. Опыт отрешенный от метафизических принципов говорит нам, что между духом и телом существует взаимная зависимость, но остающаяся всегда и у всех, тождественною. Эта безспорная зависимость, равно как и безспорная аналогия законов заставляет биологические проблемы наследственности связывать с моральными.
Прежде всего несомненен факт, что существует нравственная наследственность, как существует физическая. Указывают, что некоторые физические особенности передавались у некоторых семейств в целом ряде поколений. У Цицеронов были родинки в форме овечьяго гороха, у Лентуллов – специфические веснушки, откуда и их прозвища. В семье Роганов из поколения в поколение передавалась прядь белокурых волос на лбу, появлявшаяся в юности. Благоприятным условием для сохранения физических особенностей является заключение браков в узких границах небольшого рода. Так почти все обитатели одной долины в области верхних Альп имели по лишнему пальцу на руках и ногах. Их переселение в города лишило их потомков этой особенности. Указывают, что китайские магометане, парсы в Индии, польские евреи характеризуются некоторыми особенностями. Королевские фамилии Европы, по необходимости заключающие браки между одними и теми же семьями или по крайпей мере в ограниченном Kpyгу, дают важный материал по вопросам о наследствености. Галипп собрал более двухсот портретов, на которых были изображены члены французской, испанской, австрийской, баварской и савойской династий, между которыми брачные союзы заключались почти в каждом поколении. Семейный тип выступает. Главным образом бросаются в глаза две особенностн горбатый нос и выдайщаяся нижняя челюсть. Исследование вопроса о наследовании физических особенностей, производимое согласно принципам менделизма, привело Аперта к заключению, что физические особенности передаются лишь тогда, когда они имелись у предков и с отцовской и с материнской стороны, в противном случае они быстро исчезают.
В области нравственной указывают многочисленные примеры того, что таланты одного и того же рода проявлялись у многих членов семьи. В ряду писателей – два ІІлиния (дядя и племянник), Сенека и его племянник Лукиан, два брата Корнеля и их племяник Фонтенель, два брата Шенье, два брата Мюссе, два Александра Дюма, отец и сын. Дядя A. С. ІІушкина Василий Львович был недюжинным поэтом. В ряду натуралистов – физики Беккерели (прадед, дед, отец, сын), математики Бернулли дядя, три племянника и их сыновья), натуралисты Жоф-фруа-сент-Илеры (Этьен и Исидор, отец и сын), художники Вернэ (Карл, Иосиф и Гораций). Указывают, что наследственность талантов проявляется особенно ярко, когда брачные союзы заключаются между равно одаренными семействами. Таковы семья Дарвина и семья Гальтона, браки между которыми заключались неоднократно. Они выдвинули пять поколений выдающихся натуралистов. Эразм Дарвин, его сын – Роберт, сын Роберта – всемирно известный Чарльз, сыновья Чарльза (Фреясис, Джордж), их сыновья, из которых Джордж умер, а Леонард недавно руководилъ работами лондонскаго конгресса по Евгеник. В области музыки можно указать на семью Бахов, в области драматического искусства на семью Кокленов. ІІримеры эти можно значительно умножить.
Ηо мне думается, что на самом деле нужно резко различать наследственность способностей и наследственность таланта или гения. Относительно последняго вида наследственности я сильно сомневаюсь, что она существует, и считаю несомненным, что факт такой наследственности недоказан. Во всех приведенных примерахъ талантливых семей можно отметить, что в каждой семье выдается лишь одна крупная фигура и, не будь этой крупной фигуры, остальнея легко бы могли пройдти незамеченными. В семье Беккерелей крупным талантом является прадед Антони – Цезарь Беккерель (1788 – 1878), его потомки сохраняютъ любовь к физике и дают дельные экспериментальные исследования (главным образом в области магнитизма и электричества, отчасти – в области света), но такие исследования может производить всякий способный человек. Из Бернулл велик собственно Яков Бериулли (род. в 1654 X'.), с именем которого связаны бернуллиевы числа. Из Сент-Илеров над Исидором неизмеримо возвышается отец Этьен (1772 – 1844). Если из Дарвинов вычесть Чарльза, то останется очень немного. Затем, без всякаго труда можно насчитать десятки великих людей, потомки которых не проявляли ни малейшей даровитости и о которых история хранит гробовое молчание. Что знаем мы о детях Сократа? И вообще разве великие философы и поэты Греции вели безбрачную жизнь или были безпотомственны? Конечно, нет. Между тем в ряду учеников того или другого философа не видно его собственных детей. Конечно, здесь возможна гипотеза, что мужская талантливость парализовалась женскою бездарностью, что отрицательные свойства Ксантипиш не дали проявиться в ее детях гению Сократа. Но ведь, еще вопрос, какие были способности у Ксантиппы? Может быть она была и не глупей Аспазин, но была воспитана совершенно в иной атмосфере и на иных началах. Затем, трудно представить, чтобы великие люди Греции систематически женились на тупоумных особах. Наконец, руководясь терминологией менделистов, неужели нужно принять, что ум есть свойство рецессивное, а бездарность – доминантное. 'Гогда весь исторический процесс должен бы был прелставлять нам непрерывное торжество глупости. Но если и глупость и торжествовала иногда, как например во Франции, когда там провозгласили культ богини разума, то подобное торжество всегда являлось только спорадическим.
Думается, что ум доминантен, а глупость рецессивна. Но другое дело – талант или гений. Это – исключительные явления. ІІроизвести их путем какого-либо искусственнаго подбора и предсказать их рождение доселе оказывалось невозможным. Из безвестных семей вышли Исаак Ньютон и Луи ІІастер. Именем таланта или гения называется то, что резко возвышается над нормою. Но сочетание того, что резко возвышается над нормою, с нормальным типом не может дать промежуточного звена между гением и обыкновенным человеком. Ведь, сам гений, говоря языком менделизма, есть гений только февотипически, а генотипически он носит в себе множество свойств обыкновенных людей. Лицо, с которым соединяется гений, и фенотипически и генотипически не возвышается над нормою или по крайней мере не уклоняется от нея заметным образом. При таких условиях естественно ожидать нормальнаго потомка, По Гальтону ребенок четверт своих свойств наследует от отца. Если это правило и можетъ претендовать на приблизительную верность, тο только в тех случаях, когда оба родителя не представляют заметных уклонений от нормы.
Отмечаемый факт существования талантливых семейств на самом деле заключает в себе ту отрадную истину, что поставленный в благоприятные условия просто способный человек может стать подобным таланту. Бернулли принадлежали к семье выдающейся но своему положению. Условия воспитания Дарвинов и Гальтонов являются крайне благоприятными. ІІодсказывается вывод, что если все семьи будут в таких условиях, то все они примут вид талантливых.
Теория духовной наследственности – справедливая в пределах обычных норм – предполагает собою передачу не одних только положительных, но также и отрицательных свойств. Если передается наклонность к добродетели, то должна передаваться и наклонность к пороку. Как на иллюстрацию последняго, часто указывают на такой пример. В первое время образования английских колоний в Америке туда переселилась одна нищая, характеризовавшаяся крайнею порочностью. Пьяница, воровка, скандалистка она половину своей долгой жизни провела в тюрьме. У ней было много детей. Государственные архивы и архивы острогов и тюрем показывают, что четыре пятых из сотен ее потомков были склонны ко всевозможным преступлениям и двенадцать кончили жизнь на виселице. Из подобных примеров делают вывод, что существует наследственпость преступности, как существует наследственность гениальности. Склонный отрицать последний вид наследствеыности я еще решительнее отрицаю первый. Мир лежит во зле и нет человека, который не согрешил бы, хотя бы житие его на земле ограничивалось одним днем. Я решительно признаю свободу воли н ответствевность человека, но однако когда пред человеком предстоит дилемма: украсть или умеретъ с голоду, то ясно, что в результате мы получим или труп или вора. Потомство авантюристки, о которой расказывают американские архивы, едва ли встречало себе благоприятный прием в честном обществе. Оно естественно должно было оказаться в опозиции с этим обществом. Оно и общество были два враждебных племени. Неудивительно, что архивы общества говорят о многочисленных расправах, с членами враждебного ему рода. Об одном русском писателе, на произведениях которого лежит печать гуманности, разсказывают, что, будучи мальчиком, он обвешивал и обманывал в лавке своего отца. Правда ли это или неправда, но во всяком случае, если мальчика с самыми благородными наклонностями приучат лет с 7 – 8 к торговле по принципу ,,не надуешь, не продашь“. то он будет продавать надувая. Что по наследству передаются дурные наклонности, об этом свидетельствует все человечество, потому что в каждом человеке есть нечто дурное, но рождают ли преступники врожденных преступников? Прежде чем ответить, нужно выяснить, что значит термин „врожденный преступник“. Это – не человек, который непременно будет совершать преступление. Условия его жизни, среда могут воспрепятствовать ему делать это. Врожденный преступник, это – человек, который носит в себе неискоренимую охоту совершать преступления. В терминах менделизма, это – человек, который носит в себе неуничтожимый фактор преступности. Но существует ли такой фактор? Нет оснований предполагать его существование. О величайших преступниках мы знаем, что они кое-кого любили и кое-кому делали добро, что они имели некоторые принципы, которыми поступались. Ведь, бывали случаи и они вовсе не исключительные, что эти преступники проявляли ту любовь, о которой сказал Христос: ,,нет больте той любви, как еслн кто положит душу свою за друзей своих“ (Иоан. XV, 13).
Специфические дурные свойства – жестокость, скупость, зависть, злорадство, наклонность безкорыстно вредить ближнему – повидимому наследуются, но люди с этими свойствами в большинстве случаев бывают легальными, и самые эти свойства не представляют собою чего-то непреоборимаго, как закон роста, цвет волос или глаз, они могут усиливаться и ослабляться подъ воздействием внешних факторов.
У одного автора, который защищает теорию врожденной преступности, мы в то же время находим отрицание существования дегенератов34. ІІод дегенератами разумеются люди вырождающиеся, т. е. такие, которые хуже своих отцов и произведут потомство еще хуже себя. Наш автор – Апперт говорит, что нельзя получить в наследство от родителей такой фактор, который производил бы постепенное разрушение потомства и в каком-либо поколении привел бы его к окончательной гибели. Можно отравить зародыш алкоголем и всяким другим ядом можно передать зародышу болезнетворные микробы, все это потом на нем скажется, но все это – не факторы прогрессивной дегенерации. Явление последней на самом деле обусловливается постоянным действием какого-либо губительного фактора: неблагоприятные условия жизни, тяжелая среда, алкоголизм, пороки сказываются на каждом последующем поколении сильнее, чем на предыдущем и в конце концов приводят род к гибели. Но устраните постоянно действующий фактор, и вырождение прекратится. Так, в области физической природы. Достаточно дать телу толчок, чтобы оно задвигалось по прямому направлению и продолжало бы двигаться без конца, если бы препятствия не положили конца его движению. Но ни от какого толчка и от какой совокупности толчков тело не пойдет по линии круговой или параболической, вообще по кривой линии. Нужна постоянно действующая сила, которая постоянно отклоняла бы тело от прямого пути. В области органической и моральной для того, чтобы происходили уклонения от нормы, нужен тоже постоянно действующий фактор. В этом факте заключается многоутешительнаго. Если действительно это так, то сила начал, направляющих человечество на кривые пути, оказывается очень ограниченной. Но так как эта сила за всем тем постоянно действует в мире, то она и являет нам образы врожденных преступников, дегенератов, которых на самом деле не существует.
Теперь возникли общества и лаборатории Евгеники, поставившие своею задачею выяснение законов, обусловливающих создание хорошего потомства, добраго рода хорошо. Эти общества повидимому склонны к тому, чтобы устанавливались запрещения браков между лицами угрожающими дать человечеству духовно или физически больное потомство. Конечно, в некоторых пределах такие запрещения должны быть, но вообще они очень рискованны. Нервная развинченность может соединяться с высокою даровитостию. Гений и безумие не очень далеки друг от друга. Ньютон болел психически, Кант кончил старческим маразмом, Конт сходил с ума. Кажущаяся физическая слабость может тоже быть обманчивой. Агезилай и Суворов были хилыми ребятами. Многие рождавшиеся слабыми и имевшие повидимому плохое здоровье доживали до девятаго десятка или даже до девяноста лет, таковы Гоббес, Беккерел. Евгеника должна заботиться не столько о запрещениях, сколько об улучшении рода путем наилучшаго воспитания. Если для того, чтобы двигаться по прямому пути, достаточно одного толчка, а для того, чтобы ходить кривыми путями, нужна постоянная наличность злой силы, то очевидно, нам нужно прежде всего сосредоточить свой заботы на том, чтобы первоначальный толчок был благотворен и силен, чтобы он могуче направлял к добру. Нужно позаботиться о добром воспитании и об устранении злых влияний во время этого воспитания.
Но кроме наследственности и воспитания с нашей точки зрения существует еще свобода.
Кто верит в существование свободы, тот должен разсматривать необходимость, как неизбежное условие для проявления свободы. Определения моей свободной воли не имели бы никакого значения и смысла, если бы вокруг меня явления не совершались по законам необходимости. Если бы все вокруг меня совершалось свободыо, я не мог бы приспособить себя к среде, я немедленно погиб бы: земля могла бы уйти из-под моих ног, воздух разрелиться, дом перейдти на другое место. И если бы люди обладали неограниченной свободой, я тоже и при существовании физических законов необходимости не мог бы приспособиться к ним, как и они ко мне. И здесь требуются ограничения свободы и законосообразность, хотя и не тождественная с физической. Но за всем тем необходимость находитъ смысл своего существования лишь в том, что она обуславливает проявления свободы. Если-бы свободы не было и все совершалось бы по необходимости, то все было бы безсмысленным. Законы необходимости находят себе оправдание в законах свободы.
Но проявлению какой свободы благоприятствуют действующие в мире законы необходимости? Несомненно, необходимость можетъ содействовать проявлению и доброй воли и злой. Какой же она более содействует? Во второй половине прошлаго столетия нам, исходя из принципов дарвинизма, ответили бы, что природа равнодушна к добру и злу, и что самые понятия добра и зла условны и субъективны. Не все дарвинисты сказали бы это, но тот, кто не сказал бы этого, никаким образом не мог бы показать, каким образом вера в добро и в его торжество может соединяться с принципами дарвинизма. Теперь дарвинистическое миросозерцание отходит в область истории. Многое в природе оказалось против него. Много ударов нанес ему и менделизм. Что дадут далее работы менделистов?
Нельзя знать этого но хочется верить, что умственное движение, начало которому положил монах и в ряду продолжателей которого стоят ученые христианского образа мыслей (наприм., ботаник Викгор Грегуар), это движение во всяком случае будет содействовать установлению гармонии между нашей христианскою верою и положительным знанием, и что на вопросы о принципах природной необходимости оно будет давать ответы, поднимающие силу телеологического доказательства бытия Божия.
* * *
Донкастер–Наследственность в свете новейших изследованій. Москва. 1913.
См. об опытах Ренпо, Шрибо и Нильсона мою статью „Ботаника и дарвинизм“ (Вера и Разум. 1907, № 1).
См. описание этого опыта у Страбургера (Учебник ботаники для высших учебных заведений. Москва. 1909, стр. 280–281).
Метагенезис состоит в том, что от полового поколения организмов происходит поколение, размножающееся безполым путем, а от этого поколения происходит половое тожественное с первым.
Гетерогония обозначает происхождение от одного поколения организмов другого, на него непохожаго, потомство которого представляет возврат к 1-му поколению.
Педогенезисом называется способность полового организма размножаться безполым путем уже в личиночном (детском) состоянии.
Можно еще более упростить объяснение численного равенства полов. Представим, что самец при соединении с самкою отделяет только гаметы Ff (ясенский фактор и отсутствие женского фактора), самка. наоборот, отделяет только гаметы Mm (муясской фактор в отсутствіе мужского фактора). Предлагаемою много гипотезою прѳцпелагается, что мужчины потевціально обладают способностью производить только женщин, а женщины – только мужчин. Мне кажется, что в пользу этого предположения можно привести некоторыя данныя. Согласно ги-потезе мы получим комбинаціи FM, fm, Fm, fM. ІІервыя две безрезультатаны последние две производят женское и мужское потомство и должны производить его в равном числе.
P. К. ІІенветт-Менделизм. Москва. 1913. стр. 117.
Свою теорию Hugo de Fries наложил в обширном двухтомном труде Die Mutationstbeorie. Erster Band. 1901. Zweiter Band. 1903. ІІосле этого он не переставал развивать свои воззрения но вновь печатавшихся работах (наиболее крупная–Esp6ces et Varietes 1909). К сожалению его издания доселе не переведены на русский язык. Они могли бы и не специалистов, а просто любителей натолкнуть на поучительные наблюдения и исследования. В тульской губерни мне случайно пришлось наблюдать у чистотела (Chelidonium majus)–растения крестоцветного, как и крупка, тенденцию к образованию пятилепестковых цветов совершенно нормального типа. Это совсем не то, что представляет собою так называемое пятилепестковое „счастье» сирени. Сирени,– сростнолепестное растение, она в сущности не имеет лепесткоф, и и разрезы ее венчика дают разнообрааные отступления от освовного типа. Но эти отступления по существу и не так велики и не так часты, как наблюдавшиеся мною у чистотела. Может быть путем отбора семя можно получить пятилепестковый чистотел, который уже никак нельзя будет отнести к крестоцветным.
См. Э. Страебургера–Реальные основания учения о наследственности. СІШ. 1912. Также см. Т. Г. Моргана–Эксперимевтальная зоология. Москва. 1909.
M. E. Apert–Les Pröblemes de 1’ HeVedite (Revue seientifique. 12 Iuillet 1913).