
Естественный отбор. Дон Баттен, доктор философии (физиология растений)
Доктор Баттен является экспертом в выращивании тропических растений, в частности маша (золотистая фасоль), личи, гуавы, сметанного яблока и манго. Не будучи чуждым миру науки, Дон активно писал как для светского, так и для христианского мира, опубликовав ряд статей в научных журналах и книг. Его исследования по средовой адаптации, экофизиологии и минеральным удобрениям этих важных мировых сельскохозяйственных культур делают его отличным рефери в дискуссии по вопросу естественного отбора, поскольку в своей работе он постоянно имел дело с этой концепцией. Он рассматривает первую тему книги «Ахиллесовы пяты эволюции», имея многолетний опыт как учёного, так и креациониста.
См. creation.com/don-batten-russian
Естественный отбор: краеугольный камень дарвиновской эволюции
Полное название книги Чарльза Дарвина 1859 года отображало концепцию естественного отбора: «Происхождение видов путём естественного отбора, или сохранение благоприятных рас в борьбе за жизнь». Естественные процессы, или «Природа» [прим. пер.: англ. natural – естественный, природный], сохраняют индивидуумов, которые наиболее приспособлены к среде.
Естественный отбор на самом деле весьма очевидная и разумная идея. Создания, которые обладают особенностями (признаками), способствующими выживанию в конкретной среде, имеют больше шансов выжить, чем те, которые не обладают такими особенностями. Например, волки с меньшими ушами, более короткими лапами и толстым шерстяным покровом, имеют больше шансов выжить в Арктике, чем волки с большими ушами, длинными лапами и тонкой шерстью. Эти различия влияют на способность животных сохранять или отдавать тепло, то есть это – важные признаки для выживания в холодном или тёплом климате соответственно.
Несмотря на то, что «природа» – не живое существо, и, следовательно, не может делать никакого «выбора», естественный отбор является удобной фразой для использования при обсуждении выживания или гибели организмов и их генов с течением времени в различных средах. В 1868 году Дарвин пояснил, что у естественного отбора нет направления, нет конечной цели или задачи:
«Это сохранение отдельных разновидностей в ходе борьбы за существование за счёт того, что они обладают некоторыми преимуществами в структуре, строении или инстинкте, я назвал «естественным отбором»; а мистер Герберт Спенсер хорошо выразил эту же идею фразой «выживание наиболее приспособленных». Формулировка «естественный отбор» в некотором отношении плоха, потому что кажется, будто она подразумевает сознательный выбор; но мы забудем об этом, немного привыкнув к ней. … Для краткости я иногда говорю о естественном отборе как о разумной силе – подобно как и астрономы рассуждают о гравитации, определяющей движение планет, и как агрономы говорят о человеке как о создателе домашних видов просто потому, что он может делать выбор. И в том, и в другом случае естественный отбор не обходится без разнообразия, и это в некоторой степени зависит от окружающей среды. Я также часто персонифицирую слово «Природа», потому что мне кажется, что избежать этой двусмысленности сложно, но под природой я подразумеваю только совокупную работу и результат многих естественных законов, а под законами – только определённую последовательность событий»14.
Однако, организмам необходимо продолжать род, а не только выживать. В противном случае их признаки не будут переданы потомству. Поэтому всё, что помогает живому организму успешно размножаться (производить потомство, которое выживет и сможет продолжить род), вносит свой вклад в «приспособленность», и таким образом, в способность видов продолжать существовать в определённой среде. Насколько сильно среда влияет на приспособленность – спорный вопрос, но это было основной идеей Дарвина.
Как мы только что прочитали, Дарвин также одобрил фразу Спенсера «выживание наиболее приспособленных», но многим сегодняшним эволюционистам этот термин не нравится, потому что он склоняет людей мыслить категориями «самый большой», «самый быстрый», «самый сильный», а эти признаки не всегда увеличивают способность производить жизнеспособное потомство. «Самые приспособленные» – это, по определению, те, которые произвели наибольшее количество выживающих потомков. Тот, у кого самое большое количество детей, – выиграл! Есть некоторая путаница в этом вопросе, восходящая своим началом к самому Дарвину. Прямо перед приведённой выше выдержкой он отметил: «Как было верно сказано, вся природа находится в состоянии войны. Сильнейшие в конечном счёте одерживают победу, слабейшие проигрывают». Несмотря на эту путаницу, биологи под термином «естественный отбор» имеют в виду дифференциальное размножение. Это важное отличие.
Естественный отбор был единственным механизмом, предложенным Дарвином в «Происхождении видов» для объяснения происхождения всех разнообразных форм жизни на Земле. Всё произошло из одной первоначальной формы жизни (или из нескольких форм жизни, как допускал Дарвин). Он не знал ничего о генетике и мутациях, или об их молекулярной основе в ДНК (см. главу 2). Он предположил, что небольшие вариации происходят всегда, и те из них, которые способствуют выживанию, сохраняются, что тем самым приводит к превращению особей одного вида в особи совершенно иного вида (за достаточный период времени).
Примеры естественного отбора часто преподносятся как доказательства эволюции. Поскольку организмы часто способны адаптироваться к изменениям в окружающей среде путём естественного отбора, в таких примерах нет недостатка, и поэтому нас непрерывно бомбардируют идеей, что эволюция «происходит постоянно». Но действительно ли это эволюция?
«Специальная» и «общая» теории эволюции
Что такое эволюция? Это «изменения со временем» или «общий предок всех видов»? Пытаясь совместить обе эти идеи, теория Дарвина подразумевает формирование новых видов (видообразование), хотя на самом деле он не объяснил, как именно формируются новые виды (и то, как это происходит, до сих пор в некотором роде спорный вопрос). Об определении термина «вид» поговорим позже. Сейчас нам просто нужно понять, что видообразование просто предполагает происхождение, например, разновидности кролика, которая больше не скрещивается со своей предковой формой кроликов. Это довольно сильно отличается от представления о виде как о шаге на пути превращения микроба в человека.
Дарвин предполагал, что вариация, которую мы наблюдаем среди видов, безгранична, так что естественный отбор мог бы превратить микроба в мангуста в течение эонов времени. Он предположил, что наблюдаемые вариации между породами собак, голубей или между клювами различных видов вьюрков15 в дикой природе представляют собой тот тип изменений, которые можно практически без ограничений экстраполировать, чтобы объяснить не только разновидности вьюрков, но и само происхождение вьюрков, голубей, собак и всех остальных. Он не рассматривал эволюцию человека вплоть до опубликования 12 лет спустя своей книги «Происхождение человека и половой отбор», предположительно потому, что включение вопроса эволюции человека< в «Происхождение видов» уменьшило бы вероятность её принятия.
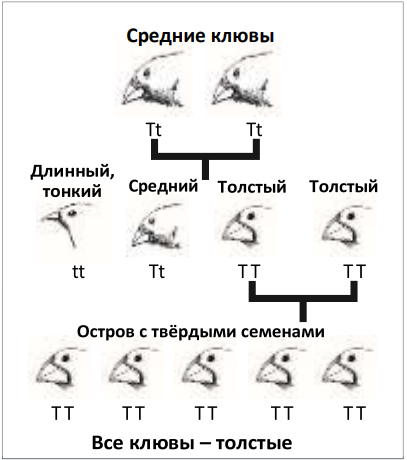
Сортировка уже существующих генов могла породить разнообразие форм клюва. Затем естественный отбор мог удалить информацию о тонких клювах
Нужно сделать огромный скачок, чтобы перейти от рассмотрения вариаций в существующем признаке (например, более короткий, тонкий, длинный, толстый клюв) к объяснению возникновения клювов, вьюрков, птиц, рептилий, млекопитающих и всего остального. Как рассмотрение вариаций собак может объяснить возникновение собак (волков)? В этом состоит фундаментальный логический разрыв, и это подчёркивает одну из важных ахиллесовых пят эволюции.
И эволюционист профессор Джеральд Керкут, известный британский авторитет в области беспозвоночных, различал «специальную теорию эволюции», СТЭ (видообразование), и «общую теорию эволюции», ОТЭ (общий предок всех живых существ). Он утверждал, что вторая является гипотетической:
Эту теорию можно назвать «Общая теория эволюции», и подтверждающие её факты нельзя назвать достаточно убедительными, чтобы мы могли считать её чем-то большим, чем рабочей гипотезой. Не ясно, имеют ли изменения, которые приводят к видообразованию, ту же природу, что и те, которые привели к развитию новых типов [крупная классификационная единица в системе живых организмов, типов существует около 80, включая микроорганизмы]. Ответ будет найден в будущей экспериментальной работе, а не в догматических утверждениях, что общая теория эволюции должна быть верной, потому что нет ничего, что может удовлетворительно её заменить.16
Кстати, Керкут включил происхождение жизни в ОТЭ. Почему же тогда столь многие не хотят включать происхождение жизни в их определение эволюции? См. главу 3.
Сейчас мы понимаем, почему простые изменения в видах (СТЭ) нельзя экстраполировать на происхождение разнообразия всего живого (ОТЭ). Тот тип наблюдаемых вариаций, который эволюционисты любят называть «эволюцией», объясняется перегруппировкой существующей генетической информации (аллелей) или случайными, и почти всегда дегенеративными, изменениями в этой существующей информации. Но эволюция от микроба к человеку требует формирования новых, сложных, переполненных информацией наборов генов, содержащих инструкции для создания, например, мышечных клеток, костей, нервов, перьев у рептилий и т.д., которых не существовало раньше. Дарвин не имел ни малейшего представления, что было бы необходимым для осуществления таких серьёзных изменений, и современная биология показала исключительную сложность, которая говорит против веры в простые изменения, накапливающиеся с течением времени.
В ноябре 1980 г. некоторые ведущие мировые эволюционные биологи провели в чикагском Музее естественной истории им. Филда конференцию, объявленную «исторической». Представляя отчёт о конференции в журнале Science, Роджер Левин написал:
Центральным вопросом чикагской конференции был такой: «Могут ли механизмы, лежащие в основе микроэволюции, быть экстраполированы для объяснения явления макроэволюции?» Рискуя серьёзно задеть позицию некоторых людей, участвовавших в обсуждении, ответ можно дать только один: «Нет».17
В этой же публикации были процитированы слова Франциско Айала, на тот момент адъюнкт-профессора генетики в Университете Калифорнии, о том, что он теперь убеждён, «что малые изменения не накапливаются».18
Тем не менее, многие эволюционисты сегодня остаются в мнимом неведении об этом. То есть они постоянно продвигают идею, что «большие изменения = маленькие изменения, умноженные на миллионы лет». Эта логическая ошибка называется подменой определений, или «замани-и- подмени» [прим. пер.: недобросовестная торговая практика привлечения покупателей рекламой одного товара по низкой цене и предложение пришедшим в магазин покупателям другого, более дорогого]. Это сродни тому, что сказать: «Раз корова может перепрыгнуть через забор, это лишь вопрос времени и практики, когда она сможет перепрыгнуть через Луну».
Некоторые преподаватели используют другие уловки подмены понятий, чтобы разоружить студентов, которые могут воспротивиться принятию общей картины эволюции (ОТЭ), рассказывая что-то вроде: «Эволюция означает изменения. Вот пример изменения, а значит, эволюция – это факт». Ещё одним неудачным определением эволюции является «изменения в частоте аллелей (генов)». Конечно, часто́ты аллелей меняются, но это не объясняет происхождение генов (вариантами коих являются аллели), а эволюция «от слизи к человеку» должна объяснять и это, а не одни только изменения в частоте существующих аллелей.
Джон Эндлер, известный эволюционист, избранный членом Национальной академии наук США, ставит точку таким образом:
Эволюцию можно определить как любое чистое направленное изменение или любое кумулятивное изменение характеристик организмов или популяций за многие поколения ... Сюда явно относятся происхождение, равно как и распространение аллелей, вариаций, характерных особенностей или признаков.19
Многие эволюционисты говорят так, будто эволюция включает в себя только последнее, и оставляют без внимания происхождение признаков. Это упущение было очевидным в работах Чарльза Дарвина и преобладает в эволюционной мысли и сегодня, но это неправильно.
Эволюция – это не просто «изменения». Это не только изменения в преобладании признаков (частоте аллелей) в популяции (СТЭ). Она также должна сопровождаться возникновением радикально новых признаков, которые являются не просто модификациями существующих (ОТЭ), и именно в этом Дарвин, а также многие после него, постоянно терпели неудачу.
Естественный отбор – это не эволюция
Многие известные эволюционисты говорят о дарвинизме (эволюции) и естественном отборе так, будто это одно и то же. Например, Ричард Докинз говорит об экспериментах, которые продемонстрировали естественный отбор на окрасе гуппи (камуфляж для защиты от хищников против яркой окраски самцов для привлечения самок) как об «эффектном примере эволюции, происходящей прямо на наших глазах».20
Доктор Джон Эндлер, процитированный выше, проводил над гуппи остроумные эксперименты. Согласился ли бы он с Докинзом, что это – «яркий пример эволюции»?21 В своей книге «Естественный отбор в дикой природе» (Natural Selection in the Wild), опубликованной в 1986 году, Эндлер чётко объяснил, почему это не одно и то же:
Естественный отбор нельзя приравнивать к эволюции, хотя эти два понятия тесно связаны.22
Естественный отбор достаточно распространён в естественных популяциях, поэтому прослеживался среди самых разнообразных организмов, и жёсткий отбор встречается не так редко, как предполагалось ранее. Поэтому, похоже, естественный отбор играет важную роль в эволюции. Однако естественный отбор не объясняет возникновение новых вариаций, он объясняет лишь процесс изменения их частоты.23
Таким образом, естественный отбор может влиять на модели возникновения комбинаций признаков, однако же он не объяснит механизмы их возникновения. Этот вопрос вскользь рассматривали в своих работах Фишер (1930), Симпсон (1944) и Ренш (1959), но с тех пор он практически не затрагивался, хотя и нуждается в дальнейших исследованиях.24
Обратите внимание, что Фишер, Симпсон и Ренш были очень известными эволюционистами. Фишер считается одним из создателей современного эволюционного синтеза (современной синтетической теории эволюции).
Что касается исследования гуппи, то хотя естественный отбор и может помочь объяснить относительную численность цветастых и менее цветастых гуппи, в зависимости от баланса между половым отбором25 (самки предпочитают цветастых партнёров) и риском быть съеденным хищником (что даёт преимущество блеклым цветам), он не объясняет происхождение цветов окраса. Даже если допустить, что некоторые мутации в уже существующем гене, отвечающем за цвет окраса, могли повлиять на внешний вид гуппи, это не объясняет происхождение самого гена. Это очень важное различие, которое нужно иметь в виду, но которое как Дарвин, так и эволюционисты после него постоянно недооценивали.
Могут ли Докинз и единомышленники действительно быть не осведомлены о факте, что естественный отбор – это не то же, что и эволюция, хотя на это открыто указывали такие знаменитые эволюционисты, одного из которых Докинз даже цитирует, когда это поддерживает его аргумент?
Следуя примеру Дарвина, эволюционисты, несмотря на утверждения об обратном, всё ещё любят говорить о естественном отборе как о созидательной силе, хотя он создать-то ничего и не может. Он может лишь устранять неприспособленных, а не создавать приспособленных. Естественный отбор – не то же самое, что эволюция. «Выживание самого приспособленного» (устранение неприспособленного) не объясняет появление приспособленного.
Естественный отбор – не открытие Дарвина
Учитывая, насколько очевидной является концепция естественного отбора, неудивительно, что эта идея возникла до Чарльза Дарвина.
Карл Линней (1707–1778), знаменитый шведский естествоиспытатель, заложивший основы используемой сегодня системы именования и классификации живых организмов, которого также называют «отцом таксономии», явно понимал принципы естественного отбора и выживания самого приспособленного. В статье о Линнее на веб-сайте Музея палеонтологии Калифорнийского университета говорится:
Линней подметил борьбу за выживание, он однажды назвал природу «колодой мясника» и «войной всех против всех». Тем не менее, он считал борьбу и конкуренцию необходимыми для поддержания равновесия в природе, частью божественного порядка.26

Слева направо: Карл Линней, Джеймс Геттон, Патрик Мэттью, Эдвард Блит. Все эти люди писали об идеях, подобных дарвиновской идее о естественном отборе, за многие годы до опубликования «Происхождения видов».
Шотландский деист Джеймс Геттон в 1794 году написал (в одной из глав своей трёхтомной неопубликованной монографии объёмом в 2000 страниц27) о собаках, которые полагались в выживании «только на быстроту ног и стремительность взгляда», в результате медленные собаки погибали, а быстрые выживали. Но если острый нюх был «более необходимым для выживания животного», то «естественной тенденцией расы, в соответствии с тем же принципом семенной вариации, должно быть изменение качеств животного и возникновение расы собак с хорошим нюхом, а не тех, кто ловит свою добычу благодаря быстроте». Он писал, что тот же «принцип вариации» должен также влиять на «каждый вид растений, будь то растущий в лесу или на лугу».28
Геттон, несомненно, пишет о естественном отборе, пусть даже точный термин не используется (сам термин был введён лишь в 1859 году, когда было опубликовано «Происхождение видов»).
Шотландско-американский врач Уильям Уэллс (1757–1817) в 1813 году описал концепцию естественного отбора. Он сказал, что в Африке некоторые обитатели
должны были быть лучше других приспособлены к перенесению заболеваний в этой местности. Следовательно, численность этой расы должна была стремительно увеличиваться, в то время как другие расы должны были сокращаться.29
Он предположил, что эта жизнеспособная раса должна была быть темнокожей и что
поскольку самая тёмная раса будет наиболее приспособлена к этому климату, то в конце концов она станет самой распространённой, если не единственной, расой в той конкретной местности, в которой она возникла.30
Шотландский садовод Патрик Мэттью (1790–1874) в 1831 году опубликовал книгу под названием «Корабельный лес и лесоводство» (On Naval Timber and Arboriculture), в приложении к которой упомянул естественный отбор. Мэтью публично заявил, что предвосхитил Чарльза Дарвина, он даже называл себя на титульных листах своих книг «первооткрывателем принципа естественного отбора».31 Дарвин отрицал, что знал о написанном Мэтью, указывая, что текст был опубликован в приложении неизвестной книги. Но даже если Дарвин и не знал об этом конкретном примере, это доказывает, что идентичные идеи циркулировали внутри британского общества викторианской эпохи и до 1859 года.
Пирсон отметил, что Уэллс, Мэтью и Дарвин все жили в университетском городе Эдинбург, «месте, знаменитом своими научными клубами и обществами», который был также родным городом Геттона.32 Действительно, было бы странно, если бы Дарвин не слышал об этих известных идеях, и возможно, что теория Дарвина, намеренно или подсознательно, развивалась в плавильном котле философских предположений, циркулировавших в его кругах общения. Суть в том, что идея естественного отбора не только не исходила от Дарвина, но и была довольно распространённой, по крайней мере среди некоторых людей того времени.
Эдвард Блит (1810–1873), вероятно, повлиял на Дарвина больше всего. Английский химик и зоолог Блит написал три наиболее значимые статьи о естественном отборе, которые публиковал в журнале The Magazine of Natural History с 1835 по 1837 гг., то есть он начал, когда Дарвин ещё был в кругосветном путешествии на корабле Бигль. Это был один из ведущих зоологических журналов того времени, и друзья Дарвина Генслоу, Дженинс и Лайель также публиковали в нём статьи.
Лорен Айзли был профессором антропологии и истории науки в Университете Пенсильвании до самой смерти. Айзли десятилетиями отслеживал происхождение идей, приписываемых Дарвину. В книге 1979 года он написал, что «ключевые догматы работы Дарвина – борьба за существование, вариация, естественный отбор и половой отбор – все полностью сформулированы в работе Блита 1835 года».33 Почему Дарвин не признал работы Блита в отношении естественного отбора? Рассел Григ предполагает:34
1. Блит был христианином и [по крайней мере на тот момент] придерживался взглядов, которые мы бы сегодня назвали «креационистскими». Например, относительно сезонных изменений в окрасе животных (скажем, заяц-беляк становится белым зимой), Блит отметил, что это «яркие примеры замысла, которые так ясно и убедительно свидетельствуют о существовании всеведущей великой Первопричины». Также он сказал, что животные «выказывают сверхчеловеческую мудрость, потому что это врождённое, а значит, привитое премудрым Творцом».35
2. Блит правильно увидел концепцию естественного отбора как механизм, с помощью которого больные, старые и непригодные удалялись из популяции; то есть как фактор сохранения, поддерживающий статус-кво – сотворённый род. Креационисты [специального сотворения], такие как Эдвард Блит [по крайней мере, на тот момент, поскольку есть данные, что после публикации «Происхождения видов» он взял на вооружение идею Дарвина о неограниченных изменениях, включая общее происхождение человека и обезьян] (и английский богослов Уильям Пейли), видели естественный отбор как процесс выбраковки; то есть как процесс выбора между несколькими признаками, все из которых должны уже существовать, прежде чем они могут быть выбраны.
Это лишь некоторые из тех, кто писал о естественном отборе до Дарвина. Мы не можем обойти вниманием Альфреда Рассела Уоллеса (1823–1913). Живя на Малайском архипелаге, он независимо разработал теорию эволюции, почти идентичную дарвиновской, включая полностью сформированную концепцию естественного отбора. В 1858 году он послал Дарвину копию своей рукописи «О тенденции разновидностей к неограниченному отклонению от первоначального типа» (On the Tendency of Varieties to Depart Indefinitely from the Original Type), в которой была изложена теория, в настоящее время известная как «дарвиновская» теория эволюции. Многие считают, что с ним очень несправедливо обошлись Дарвин и его друзья, Чарльз Лайель и Джозеф Хукер, которые боролись за то, чтобы Дарвин получил первенство в Лондонском Линнеевском обществе.

Экслибрис Эразма Дарвина с тремя гребешками на диагонали. В 1771 году он добавил слова «E conchis оmnia» (всё из ракушек), чтобы продемонстрировать свою веру в эволюцию.
Что касается эволюции (в отличие от естественного отбора), многие другие предложили идеи эволюции всех живых существ от какого-то первобытного предка (предков), в том числе ещё в дохристианские времена.36 Собственный дед Дарвина, Эразм Дарвин, антихристианин, в 1794 году опубликовал двухтомный трактат об эволюции под названием «Зоономия». Латинский девиз Е conchis omnia на его фамильном гербе переводится как «всё из раковин». Таким образом, собственный дед Дарвина не только верил в эволюцию, но и включил её в общественное лицо семьи – фамильный герб!
До Дарвина континентальная Европа видела много трудов, посвящённых эволюционным идеям, которые пытались объяснить происхождение всего без необходимости в божественном творении.37 Желание избавиться от Бога-Творца – это глубоко укоренившееся свойство человеческой натуры, которое было присуще ей и до Дарвина.
Предшественники Дарвина, в том числе Геттон и Мэттью, рассматривали естественный отбор как консервативный, или сохраняющий приспособленность, процесс, который имеет ограниченные возможности для осуществления изменений. Однако, Дарвин представил естественный отбор в качестве созидательной силы, чтобы попытаться объяснить эволюционный взгляд на происхождение всех живых существ. В этом он отличался от своих предшественников, но в этом он и ошибался, поскольку естественный отбор не является созидательным.
Естественный отбор не опровергает сотворение
Статья о Линнее на веб-сайте Музея палеонтологии Калифорнийского университета говорит:
В ранние годы Линней считал, что виды не только реальны, но и неизменны – как он написал, Unitas in omni specie ordinem ducit (неизменность вида является условием порядка [в природе]). Но Линней наблюдал, как различные виды растений могут скрещиваться, создавая формы, которые похожи на новые виды. Он отказался от концепции, что виды являются фиксированными и неизменными, и предположил, что некоторые – вероятно, большинство – видов в роду могли возникнуть после сотворения мира путём скрещивания. Пытаясь вырастить иноземные растения в Швеции, Линней также предположил, что виды растений могут изменяться в процессе акклиматизации. К концу своей жизни Линней исследовал то, что, как он думал, было случаями скрещивания между родами, и предположил, что, возможно, новые роды могут также возникнуть путём скрещивания.
Был ли Линней эволюционистом? Действительно, он отказался от своей прежней веры в неизменность видов, и это правда, что скрещивание произвело новые виды растений, а в некоторых случаях и животных. Но для Линнея процесс создания новых видов не был безграничным и бесконечным. Какие бы новые разновидности ни возникали из primae speciei, оригинальных видов в Эдемском саду, они всё равно оставались частью Божьего плана сотворения, ведь они изначально потенциально присутствовали. ...
Концепция безграничной эволюции, не обязательно управляемой Божественным Замыслом и не имеющей предопределённой цели, никогда не приходила в голову Линнею; и такая идея шокировала бы его.38
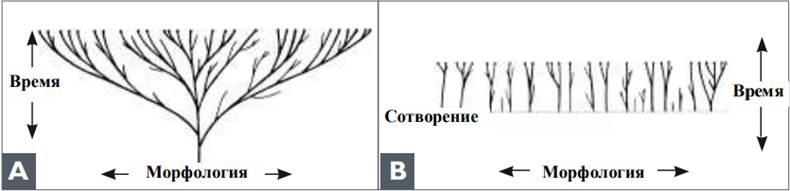
А. Оригинальное эволюционное дерево, которое предполагает, что все современные виды произошли от одного общего предка.
В. Креационистский сад. Разнообразие возникло со временем в рамках первоначальных «родов» Бытия (бараминов).
Другими словами, похоже, изначально Линней считал, что виды являются сотворёнными родами, но позже он понял, что сотворённые роды́ могут включать похожие виды и даже роды́ [прим. пер.: роды́ согласно классификации Линнея (англ. genera)]. Идея «неизменности видов» (то есть, что появление новых видов невозможно) восходит к авторитетным мыслителям древности, таким как Аристотель. Перевод Библии на латынь (Вульгата), в котором слово, обозначающее на иврите «род» (min), было переведено как «вид» (speciem/species), также подкрепил эту идею, хотя это слово переводится и как род. Поэтому, несмотря на выводы Линнея в 18-м веке, общепринятым взглядом в середине 19 века было то, что «виды» являются неизменными; что один вид может производить только такой же вид и никогда не сможет привести к возникновению другого вида. Деизм был распространён среди интеллигенции (пример – Лайель), что подразумевало за собой веру в некоторую безличную сверхъестественную «первопричину», но не Творца Спасителя, открывающегося в истории и учении Библии. Согласно этой точке зрения, «великий архитектор Вселенной» создал различные виды примерно такими же, какие они есть сегодня и примерно там, где они были обнаружены. Кроме того, официальная церковь времён Дарвина уже капитулировала перед идеей «долгих веков» и их последствием – не было глобального потопа во времена Ноя.39 Таким образом, принятие того, что можно назвать небиблейским взглядом на природу видов, не так и удивительно.
Эта «неизменность видов» подарила Дарвину прекрасного «воображаемого оппонента». Было довольно очевидно, что вьюрки на Галапагосских островах произошли от некоторых вьюрков с материка и что они постепенно породили многообразие внешне различных «видов» на островах (хотя в то время Дарвин не узнал в них вьюрков и понял их значимость, лишь когда Джон Гулд идентифицировал их как вьюрков). Вуаля! Неизменность видов опровергнута. Однако эксперименты, чтобы выяснить, могут ли скрещиваться разные виды вьюрков, никогда не проводились.
Знаменитый эскиз Дарвина «Я думаю, что» из его первого блокнота о преобразовании видов (1837 г.).
Недавние исследования показали, что скрещивание происходит даже в естественных условиях,40 то есть вьюрки Дарвина были, вероятно, даже не различными биологическими видами! А Линней признал бы их принадлежность к одному сотворённому роду.
В этих взглядах Линней предвосхитил мнение современных биологов-креационистов: организмы могут адаптироваться, формировать новые «виды». Но масштабы наблюдаемых изменений ограничены, и это не того рода изменения, которые могут объяснять происхождение принципиально разных типов организмов. То, что мы видим, – результат заложенной способности адаптироваться и диверсифицироваться в рамках сотворённых родов, благодаря чему экологические ниши Земли могли быть заполнены жизнью. Естественный отбор является составляющей этого процесса, что креационисты Линней и Блит поняли давным-давно. Так если естественный отбор является частью креационистской модели, то как он может быть опровержением концепции сотворения или доказательством эволюции как альтернативы сотворению? Если обе концепции утверждают одно и то же в данном вопросе, то это не может быть аргументом в дискуссии о сотворении и эволюции. Таким образом, дискуссия смещается с СТЭ – относительно тривиальных изменений видов, которые очевидны, просты и наблюдаемы, к ОТЭ – происхождению разнообразия всех живых существ, в которое, в свете нашего современного понимания биохимической сложности жизни, гораздо сложнее поверить, чем это было в дни Дарвина.
Что такое «вид»?
Согласно Оксфордскому словарю, биологи определяют вид как «группу живых организмов, состоящую из сходных особей, способных к обмену генами или скрещиванию». Это определение широко признано среди биологов, в том числе среди биологов-креационистов. Другими словами, если два организма могут скрещиваться между собой, они строго принадлежат к одному (биологическому) виду. Однако, из-за сложностей с проведением эксперимен тов по скрещиванию, видам часто дают названия на основании их внешнего вида (морфологии), и поэтому такая классификация не всегда соответствует барьерам для скрещивания (названий видов намного больше, чем самих биологических видов).

Сверху: род Penaeus, снизу: род Antrimpos (их возраст якобы 150 миллионов лет).
В случае с ископаемыми останками скрещивание невозможно проверить. Мышление в рамках миллионов лет и долгих эпох влияет и на то, как эволюционисты присваивают наименования ископаемым видам. Когда они находят окаменелость, которую «датируют» миллионами лет, но которая с виду очень похожа на живущий сегодня вид, их вера в общую схему эволюции заставляет их отбросить возможность, что ископаемое может принадлежать к тому же самому виду. Есть тысячи примеров «живых окаменелостей», выглядящих идентично современным видам, но им практически всегда присваивается не только другое имя вида, но и другое имя рода.41 Это неправильное именование создаёт ложное впечатление, что организмы изменились, тогда как факты говорят, что они изменились совсем незначительно или вообще не изменились. Это как нельзя лучше показывает, как исходная парадигма управляет интерпретацией «фактов».
Окаменелости, найденные на различных континентах, также часто получают различные имена, что увеличивает путаницу.
Представим, что в современном мире не существуют различные породы собак. Если бы палеонтологи нашли окаменелости собак пород немецкий дог и чихуа-хуа, интересно, какие названия они бы им дали? Практически наверняка, они дали бы им разные имена видов и, вероятно, отнесли бы к различным рода́м. А возможно, и к разным семействам? Это иллюстрирует ещё один момент: для эволюции важна не величина изменений (различий), а тип таких отличий. Хотя породы немецкий дог и чихуа-хуа выглядят совершенно по-разному, мы знаем, что эти отличия одного порядка – они заключаются в модификации существующих признаков, но не в создании чего-либо радикально нового. Ни у той, ни у другой породы нет ни перьев, ни клюва, ни каких-либо других признаков, не присущих волкоподобным. Может ли такой признак возникнуть за достаточное количество времени? Чем больше мы узнаём о сложной генетике и биохимии, стоящей за этими признаками, тем сложнее верить в эволюционное объяснение.
В Бытие 1:11 Библия говорит, что Бог создал растения: «И сказал Бог: да произрастит земля зелень, траву, сеющую семя, дерево плодовитое, приносящее по роду своему плод, в котором семя его на земле. И стало так».
Формулировка «по роду своему» повторяется 10 раз в 1-й главе Бытия применительно к созданию различных типов живых существ. Это согласуется с фундаментальным принципом биологии, который каждый наблюдает постоянно и понимает с самого детства: организмы действительно воспроизводятся по родам своим: собаки рождают собак, кошки – кошек, манговые деревья – манговые деревья и т.д. Никто никогда не наблюдал противоположного (то, как окаменелости говорят за и против изменений со временем, будет рассмотрено в главе 4), но эволюция влечёт за собой веру в то, что снова и снова один базовый тип организмов превращался во что-то совершенно иное (например, что черви со временем превратились в людей, как выразился один известный эволюционист в журнале New Scientist42). Видообразование (изменения в рамках сотворённого рода) считается доказательством более общей идеи – ОТЭ, однако, как было показано выше, видообразование является общепринятой частью креационистской модели.
Примеры скрещивания дают нам ключ к пониманию того, от каких изначально сотворённых родов произошли современные организмы. Исходя из библейского критерия родов, креационисты сделали вывод, что если два вида могут скрещиваться с настоящим оплодотворением, значит они принадлежат (произошли от) одному и тому же роду.43 Кроме того, соответственно, если один из этих видов может скрещиваться с третьим видом, то все три являются членами одного рода.44 Однако, если два существа не могут скрещиваться между собой, не обязательно следует, что они принадлежат разным родам, потому что дегенеративные изменения вследствие мутаций, таких как хромосомные перестройки, могут создать барьеры для скрещивания между особями, которые в остальных отношениях идентичны.

Косаткодельфин по кличке Кекаималу, 19-летний детёныш малой косатки и Атлантического большого дельфина (афалина), спарилась с другим дельфином, и на свет появилась самочка дельфино-косаткодельфина, которую назвали Кавили Кай (на фотографии).
В 1985 году Парк морской жизни в Гонолулу, Гавайи, сообщил о рождении детёныша от самца малой косатки (Pseudorca crassidens) и самки афалина (большого дельфина) (Tursiops truncatus).45 Рождение удивило сотрудников Парка, так как родители довольно сильно отличаются по внешнему виду. В данном случае получился гибрид между различными родами [прим. пер.: согласно классификации, созданной людьми] в одном семействе дельфиновых (Delphinidae), к которому относятся дельфины, косатки и малые (чёрные) косатки. Поскольку потомство в данном случае оказалось способно к размножению (гибридная самка впоследствии родила детёныша), эти два рода, по определению, принадлежат к одному «политипическому» биологическому виду. Другие роды в группе выглядят гораздо более похожими, чем те два, которые произвели потомство на Гавайях. Это явно свидетельствует о том, что все 12 современных родов, вероятно, произошли от одного первоначально созданного рода. Такие примеры напоминают нам, что современные таксономические классификации – это на самом деле искусственные построения, которые изначально начал Линней, но сегодня они основаны на попытке определить события ОТЭ в прошлом. В действительности она не является точной наукой, как её часто пытаются преподнести.
Креационист биолог доктор Фрэнк Марш писал об этих концепциях в 1940 году. Как сказал зоолог Уэйн Фрейр:46
Он [Марш] утверждал, и очень убедительно, что научные данные поддерживают концепцию отдельных родов, которые разнообразились и произвели все разновидности, существующие среди ископаемых и живых форм. Материал в этой книге был очень важной причиной, почему я принял концепцию «родов» как свою собственную рабочую точку зрения.
Марш придумал термин «барамин» для сотворённого рода (от евр. бара – «создать» и мин – «род») и определил критерии для различения потомков сотворённых родов. «Бараминология» – это систематическое изучение созданных родов. Например, ветеринар доктор Джин Лайтнер исследовала, принадлежат ли овцы и козы к одному и тому же барамину. Лайтнер подвела такой итог полученным результатам:
... овцы (Ovis aries) и козы (Capra hircus) принадлежат к монобарамину (группа принадлежащих к одному и тому же роду). Дополнительные данные по скрещиванию показывают, что другие виды в родах Ovis, Capra, Ammotragus, Hemitragus и, вероятно, Rupicapra, также относятся к этому монобарамину. Предполагаемый гибрид овцы и европейской косули позволяет предположить, что этот монобарамин на самом деле может включать в себя несколько семейств жвачных животных; тем не менее, прежде чем делать окончательные выводы, желательно иметь лучший задокументированный пример. Вариации, наблюдаемые в этом монобарамине, по крайней мере некоторые из которых являются адаптивными изменениями, указывают на то, что мутации и хромосомные перестройки внесли свой вклад в развитие существующих в настоящее время видов.47
Появление новых «видов» – не проблема для сотворения
Очевидно, что происхождение новых видов является частью креационистской модели. Появление новых видов действительно наблюдается. Например, «новые виды» появились в роде Дрозофил (Drosophila) – фруктовых мушек, очень популярных в студенческих генетических лабо- раториях. Довольно распространённым механизмом является инверсия – случайная хромосомная перестройка (разновидность мутаций), при которой часть хромосомы меняет своё направление. Это подавляет «генный поток» между мухами с инверсией и мухами без инверсии, таким образом, генетически разделяя их на две группы. Это позволяет им отдалиться друг от друга, даже если они всё ещё способны к гибридизации.48 Такие случаи преподносятся как «эволюция», хотя никакие новые генетические спецификации, которые были бы необходимы, чтобы изменить мушку во что-то другое, не были созданы. Всё, что произошло, – это физическая перестройка порядка генов на одной хромосоме. Это опять-таки не имеет ничего общего с метафизической верой в эволюцию от молекул к человеку – общую теорию эволюции (ОТЭ), в результате которой природные процессы произвели всё разнообразие жизни от универсального общего предка.

Обнаружено, что длина ног ящериц анолисов быстро меняется в соответствующих условиях ... наблюдаемое подтверждение библейской естественной истории.
Модель библейского сотворения включает в себя уничтожение всей дышащей воздухом, наземной жизни во время Ноева Потопа, когда лишь по паре каждого рода животных (некоторых животных по семь пар) выжили на Ноевом Ковчеге. Животные, заполняющие современные экологические ниши, произошли от тех животных, которые спаслись на Ковчеге. Поэтому мы можем заключить, что значительная диверсификация видов произошла в течение 4500 лет, прошедших с Потопа, и что видообразование может происходить иногда очень быстро. Действительно, есть множество доказательств быстрому видообразованию, которое, кажется, всегда удивляет эволюционистов из-за того, что они смотрят через призму уверенности в «долгих эпохах».49
Например, исследователи из Тринидада переместили гуппи (Poecilia reticulata) из бассейна внизу водопада, где они обитали вместе с хищниками, в бассейн над водопадом, в котором ранее не было гуппи и в котором обитал лишь один хищник, охотящийся только на маленьких гуппи, поэтому большие гуппи должны были быть в безопасности. Потомки пересаженных гуппи приспособились к новым условиям: они стали крупнее, созревали позднее и производили потомство бо́льших размеров, но в меньшем количестве.50
Скорость этих изменений удивила эволюционистов, так как согласно их концепции миллионов лет адаптация гуппи должна была происходить в течение длительного времени. Один из них сказал:
Гуппи адаптировались к своей новой среде обитания всего за четыре года – то есть скорость адаптации была в 10 тысяч – 10 млн раз быстрее, чем средние скорости адаптации, определённые по палеонтологической летописи.51
Существует множество других подобных примеров.
Процесс видообразования не подтверждает эволюцию
Ни один из наблюдавшихся примеров видообразования не подтверждает предположение о том, что оно приводит к возникновению совершенно новых организмов – новых родов. Например, в случае с упомянутыми выше исследованиями гуппи эволюционный генетик прокомментировал: «Насколько я знаю, они так и остались гуппи».52
Биологи провели всестороннее изучение цихлид – семейства рыб, обитающих на трёх континентах и популярных среди аквариумистов. Они отличаются большим разнообразием внешнего вида, и их часто приводят в качестве примера «эволюции в действии». Однако зоолог- креационист доктор Артур Джонс посвятил докторскую проблеме вариации цихлид, и его исследования укрепили его веру в библейское сотворение. Доктор Джонс пишет:
Те годы исследования были захватывающими. При всём многообразии видов я понял, что цихлиды – однозначно естественная группа, сотворённый род. Чем больше я работал с этими рыбами, тем отчётливее стал распознавать «цихлидность» и тем отчётливее эти рыбы отличались от всех «похожих» рыб, которых я изучал. Обсуждения на конференциях и изучение литературы подтвердили, что это был общий опыт специалистов в каждой области биологической систематики. Отдельные роды действительно существуют, и специалисты знают, что это так. Онтогенетические исследования показали, что огромное разнообразие цихлид (более 1 000 «видов») возникло фактически путём постоянной перестановки относительно небольшого количества состояний признаков: четыре цвета, десять или около того основных узоров окраса и так далее. Одни и те же признаки (или комбинации признаков) появлялись «случайно» по всему ареалу обитания цихлид. Схема изменений была «модульной» или «мозаичной», а эволюционные линии происхождения нигде не прослеживались. Такого рода адаптивные изменения могут происходить достаточно быстро (так как в них задействовано только то, что уже имеется в наличии), и некоторые примеры «адаптивной радиации» цихлид (в геологически «молодых» озёрах) фактически датировались (эволюционистами) в пределах не более нескольких тысяч лет. И это не очень нравится эволюционистам. Все рыбы, современные и ископаемые, принадлежат отдельным родам; «переходных форм» определённо не хватает.53
Генетические процессы, стоящие за видообразованием, включают сорти- ровку и рекомбинацию уже существующей генетической информации, как указал выше Артур Джонс. Генетические перестройки (такие как инверсии участков хромосом) могут создавать барьеры для скрещивания и способствовать видообразованию.
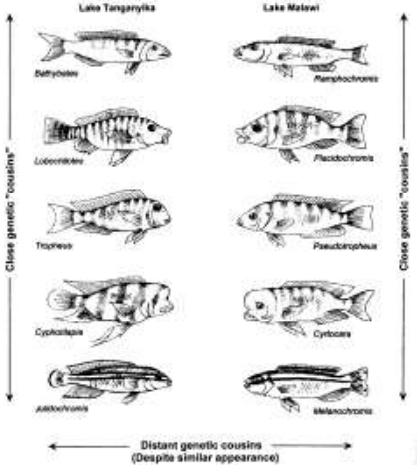
Фальк ссылается на параллельное, но независимое развитие цихлид в озёрах Танганьика и Малави в качестве доказательства эволюции. Тем не менее, это свидетельствует лишь о том, что это похожий набор варьирующихся комбинаций одних и тех же признаков из одного предкового генофонда. Kocher и др., Similar morphologies of cichlid fish in Lakes Tanganyika and Malawi are due to convergence, Molecular Phylogenetics and Evolution 2(2):158–165, 1993г.
Более того, видообразование, включающее адаптацию к конкретной экологической нише, требует устранения особей, не приспособленных к этой нише. В приведённом выше примере, волки с короткой шерстью в арктическом климате будут отсеяны, что повлечёт за собой потерю популяцией генов короткой шерсти. Поэтому процесс адаптации путём видообразования на самом деле удаляет генетическую информацию из популяции. Когда гены короткой шерсти утеряны популяцией, особи этой популяции уже не могут приспособиться к условиям жаркого климата. Они приспособлены к холоду.
Это подчёркивает проблему эволюционных взглядов на видообразование. Если адаптация путём естественного отбора постоянно удаляет генетическую информацию (вариативность), виды будут становиться всё более и более специализированными по мере того, как они всё «лучше адаптируются» к этой среде. Эта точная подстройка не позволит им пойти по обратному пути, если окружающая среда изменится. В каком-то смысле, вид бы только выиграл, оставаясь универсальным, но отбор толкает к специализации. Таким образом, с течением времени виды стали узкоспециализированными к своим нишам, из которых, возможно, уже не так легко выйти. Мы можем наблюдать это по полярным медведям: они хоть и произошли от «нормальных» медвежьих предков,54 но почувствовали бы себя не очень хорошо, если бы в Арктике потеплело и растаял лёд. Быстрое изменение многих видов путём адаптации, частично регулируемое естественным отбором, прекрасно вписывается в модель сотворения.
Никто никогда не наблюдал видообразование, которое было бы связано с появлением новых метаболических путей. Равно как никто не предложил рабочую модель таких изменений. Видообразование подразумевает пере- стройку и (или) потерю существующей генетической информации популяциями. Видообразование не подтверждает ОТЭ.
Естественный отбор не производит новой информации
Естественный отбор может работать лишь с той генетической информацией, которая определяет признаки, уже присутствующие в популяции. Он не может создавать эту информацию. Большинство эволюционистов понимают это, но они не особо стараются доводить это до широкой публики.
Современная концепция эволюции («неодарвиновский синтез») говорит, что мутации отвечают за создание новой информации (генетических инструкций, или спецификаций, для белков и т.д.). Есть много различных типов мутаций, но все из них связаны со случайными изменениями последовательностей нуклеотидов («букв») цепочки ДНК. Короче говоря, мутации – «единственная игра в городе» для создания колоссальных количеств информации во всех живых существах; инструкций по созданию, например, перьев, костей, мышц, нервов, волос, кожи, клеток крови, гемоглобина или по организации в значительной степени неизвестной последовательности событий, которые необходимы для развития эмбриона человека. Такие спецификации отсутствовали в предполагаемом первом микробе, универсальном общем предке, но мутации якобы всех их добавили, по одному незначительному изменению за раз, каждое из которых было достаточно выгодным, чтобы естественный отбор смог «увидеть» его и закрепить в популяции.
Эти последовательные мутации должны объяснять возникновение новых биохимических путей с участием многих ферментов (являющихся белками) и колоссально сложных наномашин, состоящих из многих белковых компонентов (таких как АТФ-синтаза,55 РНК-полимераза, геликаза и гираза).
Проблема в том, что даже один из компонентов этих путей или наномеханизмов не мог возникнуть случайно (посредством мутаций), не говоря уже про все компоненты, и все они нужны одновременно, чтобы многие из этих механизмов могли иметь какое-нибудь применение. Действительно, многие метаболические пути могут производить токсины, если они неполные (именно таким образом становятся токсичными некоторые патогенные бактерии, когда мутировавший ген приводит к тому, что фермент «выбивается» из метаболического пути, что влечёт за собой накопление химических веществ, которые обычно являются субстратом для недостающего фермента).
Чтобы прокариоты (такие как бактерии) превратились в эукариотов (таких как дрожжи, растения или животные), мутации должны были «изобрести» внутриклеточную транспортную систему на основе кинезина, которой обладают эукариоты и которая включает несколько интегрированных компонентов, каждый из которых необходим, чтобы она могла быть сколь-нибудь полезной.56
Это серьёзнейшие преграды для того, чтобы случайные изменения (мутации) могли изменить существующие генетические инструкции и просто непреодолимые – для создания новых.
Мутации необходимы эволюции
Сколько информации должны добавить мутации, чтобы превратить минимально возможный микроорганизм в человека? Геном простейшего микроорганизма содержит около 500000 нуклеотидов («букв»).57 Геном человека содержит около 3 миллиардов нуклеотидов. Поэтому для того чтобы первый превратился во второй, мутации должны создать почти три миллиарда нуклеотидов («букв»), содержащих информацию (инструкции). Если говорить только о белках, у простейшего микроорганизма их несколько сотен, в то время как у человека – более 100000. Более того, геном человека содержит инструкции, определяющие всю последовательность действий по развитию эмбриона (они находятся в тех 98% ДНК, которые многие биологи ошибочно назвали «мусорными» из-за своих эволюционных убеждений, см. главу 2).
Когда были открыты мутации, эволюционисты провозгласили их механизмом для создания всей необходимой информации, появившейся в ходе эволюции «из ила в учёного». Это стало главным компонентом «нового» дарвинизма, или «современного синтеза», который развился в 30–40-х годах 20-го столетия.
Ионизирующая радиация и определённые химические вещества могут способствовать мутациям, поэтому растениеводы-селекционеры с азартом принялись за неистовую «мутационную селекцию», очень надеясь на создание новых фантастических сортов и видов растений, которые будут полезны человечеству. Они увидели в этом возможность для «ускоренной эволюции». На эти программы были потрачены огромные суммы денег, но результаты оказались настолько разочаровывающими, что лишь несколько таких программ продолжают действовать в странах с развитой экономикой.
Были найдены некоторые полезные мутации, например такие, которые являются причиной карликовости (что помогает злаковым культурам не падать под весом зерна), неломких стручков (благодаря чему семена можно собрать, не рассыпая на землю), низкого содержания фитата в кукурузе (которую могут есть коровы) или цветовых вариаций декоративных цветов, например хризантем.58 Тем не менее, если рассматривать на молекулярном уровне, то каждый раз обнаруживается, что для того, чтобы произвести эти новые признаки, что-то было поломано, а не создано. Это должно быть само собой разумеющимся для карликовых растений и неломких стручков. Что касается кукурузы с низким содержанием фитата, мутации повредили метаболический путь, который производит фитат. В случае с цветами, может быть и не так очевидно, но это связано с тем, что цвет цветка определяется сочетанием различных пигментов. Мутация, которая отключает один из пигментов, приводит к новому цвету цветка, что, конечно, может придавать коммерческую ценность.59
Плодовые мушки (различные виды рода Drosophila) являются излюбленным объектом лабораторных генетических исследований, особенно в студенческих проектах. Бесчисленные эксперименты над дрозофилами с химически и радиационно стимулированными мутациями породили всевозможных дефективных мушек, но не дали никаких оснований для убеждения в том, что мутации могут создать новые генетические инструкции для преобразования этих мушек во что-нибудь иное, чем плодовые мушки.
Если где-то и должна наблюдаться эволюция, так это у микробов
Если эволюцию и можно было бы где-то наблюдать, так это у микроорганизмов. Многие виды бактерий были впервые идентифицированы и названы в конце 19 века, и они по сей день имеют точно такие же идентифицирующие признаки.
Одна из тенденций у бактерий, идущая вразрез с «направленной вверх» эволюцией, которая подразумевает появление новых генов, – это тенденция к удалению генов.60 Прежде всего это относится к облигатным паразитам и культивируемым в питательной среде микробам, когда без многих генов первоначальной формы уже можно обойтись, и они, как правило, утрачиваются путём удаления.
В 1988 году доктор Ричард Ленски из Университета штата Мичиган посеял 12 культур кишечной палочки (E. coli) и выращивал их в своей лаборатории, поколение за поколением, в течение двадцати лет (он заслуживает похвалы за настойчивость). Питательная среда содержала некоторое количество глюкозы, но в основном – цитрат, поэтому, как только микробы употребляли всю глюкозу, они могли продолжать расти, только развив какой-то способ потребления цитрата. Ленски ожидал быстро увидеть эволюцию в действии. Это было закономерное ожидание для тех, кто верит в эволюцию, потому что бактерии быстро размножаются и могут образовывать огромные популяции, как в данном случае. Также они могут претерпевать более высокие скорости мутации, чем организмы с гораздо более крупными геномами, как, например, люди и другие позвоночные. Всё это укрепляет уверенность неодарвинистов, что таким образом можно увидеть много примеров эволюции, происходящей в режиме реального времени (а не просто представлять, как это всё происходило в ненаблюдаемом прошлом). С учётом короткого времени генерации бактерий за 20 лет сменилось около 44000 поколений, что эквивалентно нескольким миллионам лет смены поколений человеческой популяции. Но у человека эволюционных возможностей было бы гораздо, гораздо меньше из-за: относительно небольшого размера популяции, что ограничивает количество мутационных возможностей, гораздо большего и намного более сложного генома, который не может выдерживать аналогичные темпы мутаций, не приводя к катастрофе ошибок (то есть исчезновению), а также полового размножения, из-за которого существует значительная вероятность не передать потомству выгодную мутацию.
После нескольких безрезультативных лет Ленски, похоже, отказался от идеи увидеть «эволюцию в лаборатории» и обратился к компьютерному моделированию эволюции с помощью компьютерной программы под названием Avida. Действительно, у Ленски были объективные основания потерять надежду. Он рассчитал, что все возможные простые мутации должны произойти по несколько раз, но не возникло ни одного даже простого адаптивного признака. Но позже с большой помпой было объявлено, что один из их 12 штаммов приобрёл способность использовать цитрат в 31500-м поколении.
Теперь кишечная палочка уже может употреблять цитрат в анаэробных условиях. И всё, что должно произойти для того, чтобы она могла использовать его в аэробных условиях, – это поломать механизм, который подавляет поглощение цитрата в присутствии кислорода. Я предположил это в 2008 году.61 Намного легче сломать что-нибудь, чем создать, и эволюция, как мы видели, имеет весьма скудные достижения в создании чего-либо. Лаборатория Ленски впоследствии нашла мутации, ответственные за эту новую способность, о чём было опубликовано в 2012 году.62 Цитрат доставляется в клетку с помощью белка-транспортёра. Этот белок определяется геном транспортёра цитрата, citT, который обычно выключен в присутствии кислорода. В непосредственной близости от гена citT находятся гены, имеющие промотор, который включает эти гены в присутствии кислорода. Одна мутация привела к дублированию промотора в место, которое включает в ген citT, в результате белок транспортёра цитрата теперь производится в присутствии кислорода. Дальнейшие мутации дублировали citT, то есть производилось больше транспортеров цитрата, так что больше цитрата поглощалось.
Как я и предполагал, будучи биологом-креационистом, что-то было повреждено – механизм, который подавлял citT в присутствии кислорода. Мутации не создали новых генов или даже нового промотора; они просто скопировали и вставили то, что уже было в геноме бактерии, произведя бактерию, которая больше не может выключить ген citT в присутствии кислорода (обратите внимание, что вне искусственной среды лаборатории эти бактерии были бы неадаптированными, потому что производить транспортные белки, когда они не нужны, было бы расточительно).
Это согласуется с анализом молекулярного биолога доктора Майкла Бехе, который рассматривал другого микроба с огромными популяциями организмов – малярийного паразита Plasmodium.63 Этот микроб развил сопротивляемость к различным противомалярийным препаратам, а люди развили некоторую меру сопротивляемости к нему (например, серповидно-клеточная анемия и талассемия). Бехе показывает, что все случаи адаптации, как у Plasmodium, так и у людей, происходят из-за повреждения чего-то, а не в результате создания новых сложных функций. Например, сопротивляемость к хлорохину у Plasmodium возникает из-за неисправности в транспортном белке, который доставляет яд в вакуоли организма. Бехе сравнивает борьбу с позиционной войной, в которой обороняющиеся войска уничтожают свой собственный мост или взрывают дорогу, чтобы препятствовать продвижению противника. Это на самом деле не гонка вооружений, потому что в гонке вооружений противостоящие силы изобретают новое оружие. Природные процессы («эволюция»), действующие в Plasmodium и в людях, не изобрели новое оружие.
Бехе также изучал сопротивляемость к пириметамину в Plasmodium, сопротивляемость к ДДТ у комаров и сопротивляемость к варфарину у крыс. В каждом из этих случаев что-то было повреждено в результате мутаций, что привело к возникновению сопротивляемости.
Существование базы данных мутаций, которые вызывают болезни человека, подчёркивает губительное влияние, которое оказывают мутации. Например, в онлайн-версии базы данных «Менделевское наследование у человека»64 по состоянию на октябрь 2011 года были перечислены 2 665 генов с одной или более мутациями, которые, как известно, вызывают заболевания. Органы здравоохранения обеспокоены воздействием на людей мутагенов и ионизирующего излучения, потому что мутации, которые они вызывают, приводят к заболеваниям, а не создают «людей икс» (фантастических суперменов, чьи новые способности и силы якобы появились благодаря мутациям).
Результаты мутационной селекции и мутации в микроорганизмах ещё раз подчёркивают, что случайные изменения (мутации) не создают генетических спецификаций, необходимых, чтобы превратить простые организмы в более сложные с новыми признаками, для которых требуются дополнительные ДНК-инструкции (совершенно новые гены и их системы контроля).
Тем не менее, мутации того или иного типа являются единственным механизмом, с помощью которого эволюционисты объясняют почти непостижимые количества генетической информации в биосфере.
Изменения с течением времени? Да. Изменения, которые позволили микробам стать микробиологами? Нет! Это просто бессмысленно.
Естественный отбор – сохраняющий процесс
Дарвин считал, что естественный отбор может выявить даже наименьшие вариации. Но это было наивно. На самом деле естественный отбор может отбирать лишь сильно выраженные эффекты, то есть экстремальные изменения, такие как что-то, что приводит к смерти или предотвращает смерть. Кимура понял, что эффект большинства мутаций слишком слаб, чтобы естественный отбор мог с ними работать. Существует диапазон мутаций, которые невидимы для естественного отбора. Такие мутации находятся в так называемом «диапазоне Кимуры».

Диапазон Кимуры: большинство мутаций вредны (слева от вертикальной линии), но лишь незначительно (и поэтому они скапливаются вдоль нулевой оси). Полезные мутации, как считается, очень редки (они образуют небольшой изгиб справа от нулевой оси). Но любые мутации с небольшим эффектом (рядом с линией) не видны естественному отбору и будут сохраняться. Они эффективно скрыты от отбора.
Другой эволюционист, Джон Холдейн, определил в 1950-х, что признаки должны обладать селективным преимуществом хотя бы в 10%, чтобы естественный отбор имел реальный шанс на закрепление их в популяции (10%-ное селективное преимущество означает, что организмы, имеющие признак, производят на 10% больше выживших потомков, чем те, которые его не имеют).65
Эволюционисты предположили, на основе популяционной генетики и их гипотез о временных рамках и процессе эволюции, что частота мутаций в организмах является довольно низкой, порядка 1 на человека за поколение или меньше. Признавая вредность большинства мутаций, они должны были предположить, что их было немного, иначе естественный отбор не смог бы избавиться от преобладания плохих мутаций; наступила бы мутационная катастрофа, особенно в течение предполагаемых ими временных эпох длиной в миллионы лет.
Однако, в последние годы частота мутаций была измерена, и она как минимум в 50 раз выше, чем предполагалось на основе эволюционной идеологии. Это является огромной проблемой для всей идеи. Доктор Джон Сэнфорд, вышедший на пенсию генетик Корнельского университета (в настоящее время почётный адъюнкт-профессор) и изобретатель генной пушки,66 показал, что эта высокая частота мутаций, в сочетании с тем, что большинство мутаций немного вредны (попадают в диапазон Кимуры), означает, что эти слегка вредные мутации являются невидимыми для естественного отбора и накапливаются в организме человека и других живых организмов. Этот процесс неизбежен, и это разрушает нас, а не создаёт. Мы движемся к вымиранию, как и любой другой сложный организм.67
Сэнфорд спрашивает: если большинство несущих информацию нуклеотидов («букв» ДНК) по отдельности вносят бесконечно малый вклад в геном, то как они туда попали и как они остаются там в течение «долгих эпох»? Естественный отбор не может видеть их по отдельности; он может «видеть» только общую приспособленность (способность выживать и размножаться), и вклад любого отдельного нуклеотида, как правило, настолько мал, что он невидим; он похоронен в «шуме» всех остальных нуклеотидов. Доктор Сэнфорд резюмировал проблемы для эволюции:68
1) мутации возникают быстрее, чем отбор может устранить их;
2) мутации в подавляющем большинстве являются слишком трудноразличимыми, чтобы их возможно было «отобрать»;
3) биологический шум и «выживание самого удачливого» сводят на нет отбор;
4) плохие мутации часто физически связаны с хорошими мутациями, так что они не могут быть разделены при наследовании (чтобы избавиться от плохих и сохранить хорошие). В результате все геномы высших организмов должны неизбежно вырождаться.
Знающие об этих проблемах эволюционисты предложили в качестве решения «синергетический эпистаз» (где последствия нескольких мутаций, происходящих вместе, якобы сильнее, чем их сумма).69 Однако Сэнфорд показал, что это лишь усугубило бы проблему. Доктор Томоко Охта, эволюционистка и главный воспитанник Кимуры, вместе с которым они опубликовали много работ, известна как «королева популяционной генетики». Она является почётным членом Американской академии наук.
Один из сотрудников Сэнфорда спросил её о синергетическом эпистазе, и она согласилась, что он лишь усугубил бы проблему.70 Сэнфорд и его коллеги также провели численное моделирование, используя сложную модель популяционной генетики под названием «Счетовод Менделя»,71 чтобы показать это.
Процесс, который неуклонно ведёт к деградации генома (увеличивает «генетическую энтропию»), не может породить более совершенный организм в долгосрочной перспективе. Анализ Сэнфорда является разрушительным для эволюционной парадигмы. Трудно понять, как её можно продолжать отстаивать в свете этого критического анализа. И поскольку об этом говорит такой известный генетик, то к этому нельзя относиться несерьёзно.
Тем не менее, эволюционисты постоянно приводят примеры «эволюции» с помощью адаптивных мутаций, пытаясь убедить нас, что она действительно работает. Однако они приводят примеры следующего рода: потеря зрения пещерными рыбами72 и пещерными саламандрами,73 потеря функциональных крыльев жуками на ветреном острове,74 потеря контроля над производством фермента или дефектный канал всасывания, обеспечивший устойчивость к антибиотикам,75 и дефектный ген рыбы атлантический томкод, который помогает ей выжить в водах, загрязнён- ных полихлорированными бифенилами (ПХБ).76 То есть нам дают «поломанные» организмы в качестве примеров адаптивных мутаций и естественного отбора.
Колюшки
Современная «икона эволюции», рыба колюшка (различные виды рода Gasterosteus), является ещё одним примером естественного отбора и, возможно, даже адаптивных (полезных) мутаций, но опять-таки показывает, что эти процессы не подтверждают эволюцию «от рыбы до рыбака» (ОТЭ).
Колюшки бывают морские (живущие в солёной воде) и пресноводные (обитающие в озёрах). У морских имеются заметные шипы-колючки на теле и многочисленные защитные костяные пластинки. Они помогают рыбе защищаться от хищников. Пресноводная разновидность демонстрирует большое разнообразие в морфологии, но эти рыбы, как правило, имеют более короткие спинные и брюшные колючки и значительно меньше костяных пластинок; у некоторых брюшные колючки или костяные пластинки вообще отсутствуют.77
Считается, что пресноводная форма «эволюционировала» из морской формы. Идея о том, что пресноводный вид произошёл от морского, имеет под собой основания и прекрасно вписывается в модель сотворения с послепотопной колонизацией и диверсификацией.
Морские формы мигрируют вверх по течению к пресноводным озёрам на нерест, и различные формы могут скрещиваться, что указывает на то, что они принадлежат к одному созданному роду (барамину).
Биологи обнаружили, что когда морские колюшки попадают в пресноводные озёра, спустя несколько лет количество защитных пластинок и размер колючек уменьшаются. Ряд факторов делают пластинки и колючки «неполезными» в озёрах: отсутствие крупных хищников, «затраты» на создание пластинок в бедной на кальций озёрной воде и наличие на дне озера хищных личинок стрекозы, которые используют брюшные колючки, чтобы зацепиться за колюшку и плавать с ней. Колюшки с меньшим количеством пластинок и лишённые брюшных колючек явно более «приспособлены к выживанию» в условиях озера, и естественный отбор увеличивает количество рыб с меньшим количеством пластинок и без колючек.

Что стоит за этими изменениями? Была ли изобретена «эволюцией» какая-то новая функция? Генетики обнаружили мутировавший генетический переключатель, который влияет на экспрессию гена под названием Pitx. В брюшной области повреждённый генетический переключатель предотвращает образование колючек. В других местах он влияет на «целый ряд костяных признаков»,78 не только на внешние костные пластинки, но и на форму челюсти и костей, связанных с защитой жабр. Другими словами, пресноводные формы колюшек произошли путём повреждения генетического переключателя, в результате чего определённые признаки были отключены.
Даже ведущий сторонник всего эволюционного, доктор Джерри Койн (Университет Чикаго), говоря об этих мутациях в генетических переключателях колюшки, чистосердечно признался: «Эти примеры представляют собой утрату признаков, а не появление эволюционных новшеств».79 И всё же эти различия до сих пор выставляют напоказ как «доказательства эволюции». В 2009 году, объявленном «Годом Дарвина», журнал Nature удостоил колюшек места среди «15 эволюционных жемчужин». Один известный эволюционист, Шон Кэрролл, назвал их «одним из самых убедительных конкретных примеров эволюции».80
Мы видим ещё один хороший пример естественного отбора, адаптивной мутации, и даже видообразования, но никакого подтверждения тому, что колючки и защитные пластины могли возникнуть изначально, путём мутаций и естественного отбора.
Даже мутировавший генетический переключатель, пусть и слабо распространённый, уже присутствовал в морской форме колюшек.
Также наблюдалось обратное описанному выше: в очищенных от загряз- нения частях озера Вашингтон (недалеко от Сиэтла), где видимость возросла настолько, что колюшки со слабой «бронёй» стали более уязвимы для хищников – лосося Кларка, количество полностью бронированных колюшек увеличилось с 6% до 49% в течение пяти десятилетий. Это превозносилось как «быстрая, обратная эволюция». Однако, опять же, ничего нового «эволюцией» придумано не было. Гены полной брони уже присутствовали в популяции (первоначально 6%); естественный отбор просто увеличил частоту этих генов.81 Это не более чем обычный трюк подмены определений: «эволюция – это просто изменение частоты генов».
В этом нет ничего, что подтверждает общую теорию эволюции (ОТЭ), которой необходимо изобретение наборов новых генов, а не повреждение существующего генетического переключателя (в любом случае, мутировавший переключатель уже существовал) или изменение частоты существующих генов.
Генетические переключатели созданы для облегчения адаптации?
Ветеринар-исследователь креационист доктор Джин Лайтнер предположила, что генетические переключатели, подобные тому, который действует у колюшек, могли быть умышленно созданы для упрощения адаптации, с заложенной склонностью к мутациям, но без отрицательного влияния на другие признаки. Она обнаружила, что определённый белок действует как многофункциональный переключатель, создающий широкое разнообразие окрасов шерсти у животных. Этот белок влияет только на цвет шерсти, поэтому мутации переносятся хорошо. Когда этот белок функционирует нормально, он может включить производство тёмного пигмента под названием эумеланин. Также вырабатывается более светлый пигмент, имеющий цвет от жёлтого до красного. Мутации могут поломать переключатель так, что он уже не стимулирует выработку тёмного пигмента. Это приводит, например, к желтовато-коричневому (светлому) окрасу собак породы ретривер. И наоборот, «переключатель, который заклинило во включённом положении», чрезмерно стимулирует выработку эумеланина, и это приводит к очень чёрному цвету.
Доктор Лайтнер отметила:
Создатель вполне мог бы разработать этот специальный ген-переключатель с возможностью варьироваться таким образом, чтобы создать разнообразие цветовых узоров.82
Многие такие генетические переключатели, вероятно, существуют для того, чтобы сделать возможными изменения и адаптацию. Тем не менее, эволюционисты не могут предложить механизм, объясняющий их существование. Все их «доказательства эволюции» – не что иное, как примеры модификации существующих переключателей.
Когда говорят о «мутациях с обретением функций», это звучит так, будто они включают в себя мутации, которые добавляют новые функции/признаки/гены организмам, а не просто что-то ломают. Тем не менее, эволюционисты определили «мутации с обретением функций» как «мутации, которые изменяют или повышают активность белка». Доктор Лайтнер приводит примеры.83 Один из них связан с регулированием выработки гормона щитовидной железы. В данном случае рецептор, который регулирует выработку гормона, может мутировать так, что «заклинит» и не будет реагировать на гормон, стимулирующий щитовидную железу (тиреотропин), из-за чего та будет производить чрезмерное количество гормона щитовидной железы. «Повышенная активность белка»? Да, но избыток гормона щитовидной железы приводит к заболеванию.
Лайтнер пишет:
Несмотря на обманчивую формулировку в определении «приобретения функций», здесь нет увеличения количества информации или усовершенствования метаболических путей. Без механизма развития таких путей эволюция – не более чем миф.
Эпигенетика мешает естественному отбору
Недавнее открытие эпигенетики добавило ещё одну проблему для естественного отбора.
Эпигенетика – это модифицирование активности генов, главным образом с помощью прикрепления метильных групп (–СН3) к определённым нуклеотидам (к цитозину) в ДНК. Этот процесс называется метилированием. Объёмистая метильная группа блокирует механизм транскрипции (РНК-полимеразу), так что метилированный ген не производит никакого белкового продукта (он «подавлен»).
Окружающая среда влияет на метилирование. Появляется всё больше подтверждений тому, что метилированные гены могут передаваться от одного поколения к следующему.84 И метилирование можно обратить вспять. Примеры эпигенетического наследования включают в себя количество лепестков у растений и окрас шерсти у мышей. У мышей при определённых условиях рацион матери влияет на цвет шерсти её потомства. Цвет шерсти может также передаваться к мышам-внукам, но эффект сходит на нет в течение поколений, если рацион изменяется. Устранение фактора окружающей среды позволяет паттернам метилирования вернуться в исходное состояние.85
Такие эпигенетические эффекты ещё более затрудняют работу естественного отбора, потому что у нас появляется передвигающаяся мишень. Как объясняет доктор Роберт Картер:
Например, скажем, рацион мышей вызывает эпигенетические изменения у потомства, в результате которых мыши приобретают цвет шерсти, который обеспечивает хорошую маскировку. Мышь без такого окраса шерсти может оказаться в невыгодном положении. Однако между самими генами, которые определяют цвет шерсти, нет никакого различия. Мыши имеют разные цвета в зависимости от того, что они ели. Будут устранены и некоторые особи, у которых не было генетических недостатков, и когда эффект сходит на нет, весь этот отбор оказывается впустую.86
Эпигенетика добавляет ещё больше «шума» в то, что «слышит» естественный отбор. Это, в сочетании с невидимостью многих слегка вредных мутаций и невидимостью почти всех редких выгодных мутаций, заставляет удивляться тому, что вообще есть какие-либо примеры работающего естественного отбора.
Естественный отбор слишком медленен
Знаменитый эволюционный генетик Джон Холдейн (1892–1964) был одним из основателей научной области, известной сегодня как популяционная генетика. Холдейн обнаружил серьёзную проблему для эволюционной генетики и описал её в ключевой статье, опубликованной в 1957 году, – «стоимость замены».87 Когда в популяции возникает полезная мутация, её частота должна увеличиться, чтобы популяция эволюционировала (если мутация осталась у единственной особи, эволюционный прогресс не происходит; это совершенно очевидно). Другими словами, она должна заменить (заместить) немутировавшие гены в популяции. Но скорость, с которой это может происходить, ограничена. Главным фактором, ограничивающим скорость замещения, является коэффициент воспроизводства данного вида. Для человекоподобных существ с длительностью поколения около 20 лет и низким коэффициентом воспроизводства скорость распространения мутации в популяции будет крайне низкой.
Представим популяцию, состоящую из 100000 обезьян, предполагаемых предков человека. Предположим, что самец и самка оба наследовали мутацию, настолько полезную, что они пережили всех остальных. Вся остальная популяция вымерла – все 99998 особей. А затем выжившая пара произвела достаточное количество потомства, чтобы восполнить популяцию до прежнего количества за одно поколение! И то же самое повторялось каждое поколение (каждые 20 лет) в течение 10 миллионов лет, то есть более, чем прошло времени от предполагаемого ближайшего общего предка людей и шимпанзе. Это означало бы, что 500000 (10000000/20) полезных мутаций могли быть добавлены в популяцию. Даже согласно такому совершенно нереалистичному сценарию, максимально благоприятному для эволюционного прогресса, могло быть образовано лишь 0,02% генома человека. С учётом того, что различие между ДНК человека и шимпанзе, нашего предполагаемого ближайшего ныне живущего родственника, составляет по меньшей мере 5%, или 150 миллионов нуклеотидных оснований88 («букв»), у теории эволюции серьёзные проблемы с объяснением происхождения генетической информации в таком создании, как человек. Именно отсюда произошла идея «мусорной ДНК»; поскольку эволюционисты понимают, что естественный отбор не мог создать такое количество ДНК, они посчитали, что подавляющая её часть должна быть нефункциональной (подробнее о мусорной ДНК см. в следующей главе).
Однако, при более реалистичных оценках коэффициентов приспособленности/отбора и коэффициента воспроизводства количество полезных мутаций, которые могут успеть нако- питься, стремительно падает. Согласно вычислениям Холдейна, не более 1667 полезных замещений могло произойти за предполагаемые 10 млн. лет, прошедших с последнего общего предка обезьян и людей. Это всего лишь в среднем одно замещение за 300 поколений. Происхождение всего того, что уникально отличает нас как людей, должно быть объяснено в рамках этого ограничения.
В том смысле, в котором это слово использовал Холдейн (в популяционной генетике), замещение является одиночным мутационным событием, которое распространяется по всей популяции (становится «фиксированным»). Это может быть дублирование гена, хромосомная инверсия либо же вставка, замещение или удаление единичного нуклеотида. Биологи обнаружили, что подавляющее большинство замещений на самом деле являются однонуклеотидными, поэтому предел Холдейна накладывает серьёзные ограничения на возможности эволюции, потому что 1667 однонуклеотидных замещений – это менее одного гена среднего размера.
Следует также отметить, что Холдейн сделал ряд упрощающих допущений, которые фактически оптимизируют количество возможных замещений. Таким образом, в действительности число возможных замещений намного меньше, чем рассчитал Холдейн.
Анализ Холдейна показывает, что естественный отбор мутаций не может объяснить геномы таких организмов, как человек; он происходит слишком медленно. Эволюционисты сделали всё возможное, чтобы попытаться игнорировать эту проблему, но Уолтер РеМайн недавно попытался проломить лёд в этом вопросе с новаторским анализом проблемы, а также прояснить некоторую путаницу, которая была у Холдейна по этому вопросу.89
Естественный отбор, если он и работает, говорит в пользу креационистской позиции
Работает ли естественный отбор вообще? Что ж, в некоторых ситуациях, похоже, что да, как мы это видели на некоторых примерах. Но его роль преувеличена. Некоторые классические примеры естественного отбора, такие как хвост павлина, который, по предположению Дарвина, появился в результате того, что самки предпочитают самцов с эффектными хвостами («половой отбор»), оказались безосновательными домыслами.90 И опять-таки, даже если бы пояснение отбора прошло экспериментальную проверку, оно бы объясняло не возникновение хвоста,91 а всего лишь, возможно, его сохранение, поскольку естественный отбор является консервативной силой, каким его первоначально представлял себе Эдвард Блит, креационист специального сотворения 19-го века.
Куда это ведёт?
Мутации и естественный отбор явно не подходят на роль механизма общей теории эволюции. Современный прорыв в области молекулярной биологии стал всего лишь ещё одним аргументом против эволюции как разумного объяснения разнообразия жизни на Земле. В следующей главе будет рассмотрено то, что стоит за эволюционной теорией, – область генетики, и то, как современные открытия в области генетики делают идею эволюции (ОТЭ) совсем несостоятельной.
* * *
Darwin, C.R. The variation of animals and plants under domestication, 1-е изд., т. 1, выпуск 1, John Murray, Лондон, Великобритания, стр. 6, 1868 г.
Дарвин изучал этих вьюрков, когда был на Галапагосе. См. Wieland, C., Darwin’s finches, Creation 14(3):22–23, 1992 г.; creation.com/darwins-finches.
Kerkut, G.A., Implications of Evolution, Pergamon, Оксфорд, Великобритания, стр. 157, 1960 г.
Lewin, R., Evolutionary theory under fire, Science 210(4472):883–887, 1980 г.
Lewin, ссылка 4, стр. 884.
Endler, J. A., Natural Selection in the Wild, Princeton University Press, Нью-Джерси, США, стр. 5, 1986 г.
Dawkins, R., The Greatest Show on Earth, Free Press, Нью-Йорк, стр. 139, 2009 г. См. также Sarfati, J., Dawkins playing bait and switch with guppy selection, февраль 2010 г.; creation.com/dawkins-guppy.
См. Catchpoole, D., Defining terms, январь 2011 г.; creation.com/defining-terms
Endler, J.A., ссылка 6, стр. 8.
Endler, J.A., ссылка 6, стр. 245.
Endler, J.A., ссылка 6, стр. 246.
Половой отбор – это особая форма естественного отбора, в которой самец или самка некоторого вида оказывает предпочтение в спаривания партнёру, обладающему определёнными признаками (например, цветом).
Waggoner, B., Carl Linnaeus (1707–1778), пересмотрено и обновлено в 2000 г.; ucmp.berkeley.edu
Pearson, P., Nature 425(6959):665, 2003 г. В своём обзоре Пирсон говорит, что Геттон «использует механизм отбора, чтобы объяснить происхождение разновидностей в природе», хотя «он определённо отвергал идею эволюции между видами как «романтическую фантазию"».
Цит. по Pearson, P., ссылка 14.
Цит. по Gould, S.J., Natural selection as a creative force, The Structure of Evolutionary Theory, Belknap Press of Harvard University, Кембридж, Массачусетс, США, стр. 138, 2002 г.
Цит. по Gould, S.J., ссылка 16.
Pearson, P., ссылка 14.
Pearson, P., ссылка 14.
Eiseley, L., Darwin and the Mysterious Mr X, E.P. Dutton, Нью-Йорк, США, стр. 55, 1979 г.
Grigg, R., Darwin’s illegitimate brainchild, Creation 26(2):39–41, 2004 г.; creation.com/brainchild.
Blyth, E., цит. по Eiseley, L., ссылка 20, стр. 108.
Bergman, J., Evolutionary naturalism: an ancient idea, J. Creation 15(2):77–80, 2001 г.; creation.com/ naturalism-old.
Bergman, J., Did Darwin plagiarize his evolution theory? J. Creation 16(3):58–63, 2002 г.; creation.com/ darwin-plagiarism.
Waggoner, B., ссылка 13.
Например, церковники, которые были наставниками молодого Дарвина в Кембриджском университете, см. Grigg, R., Darwin’s mentors, Creation 32(1):50–52, 2009 г.; creation.com/ clergy-mentor-darwin.
Wieland, C., Book review: The Beak of the Finch; Evolution in Real Time, J. Creation 9(1):21–24, 1995 г.; creation.com/beak_finch.
Каталог с многими подобными примерами из мезозойских пород (т.н. «эра динозавров»), см. Werner, C., Evolution: the Grand Experiment Vol. 2– Living Fossils, New Leaf Press, Грин-Форест, Арканзас, США, 2009 г.
Британский палеонтолог Саймон Конвей Моррис заявил, что «однажды мы были червями», см. New Scientist 179(2406):34, 2003 г.
Marsh, F.L., Variation and Fixity in Nature, Pacific Press, Маунтин-Вью, Калифорния, США, стр. 37, 1976 г.
Scherer, S., Basic Types of Life, в Scherer, S. (ред.), Typen des Lebens, Pascal-Verlag, Берлин, стр. 197, 1993 г.; Dembski, W.A., Mere Creation, InterVarsity Press, Даунерс-Грув, Иллинойс, США, гл. 8, 1998 г.
Batten, D., Ligers and wholphins? What next? Creation 22(3):28–33, 2000 г.; creation.com/liger.
Frair, W., My experiences as a creationist student in zoology departments of several universities, март 2011 г.; creation.com/frair.
Lightner, J.K., Identification of species within the sheep-goat kind (Tsoan monobaramin), J. Creation 20(3):61–65, 2006 г.; creation.com/sheep-kind.
Stevison, L.S., Hoehn, K.B. и Noor, M.A.F., Effects of inversions on within-and between-species recombination and divergence, Genome Biol. Evol. 3:830–841, 2011 г.
Catchpoole, D. и Wieland, C., Speedy species surprise, Creation 23(2):13–15, 2001 г.; creation.com/ speedy-species-surprise.
Reznick, D.N. и др., Evaluation of the rate of evolution in natural populations of guppies (Poecilia reticulata), Science 275(5308):1934–1937, 1997 г.
Morell, V., Predator-free guppies take an evolutionary leap forward, Science 275(5308):1880, 1997 г.
Morell, V., ссылка 38.
Arthur Jones, Biology, в Ashton, J. (ред.), In Six Days – Why 50 Scientists Choose to Believe in Creation, Master Books, Грин-Форест, Арканзас, США, 2001 г., стр. 241–248; creation.com/jones. Хорошая иллюстрация разнообразия цихлид в озёрах Африки есть в: Kocher, T.D. и др., Similar morphologies of cichlid fish in Lakes Tanganyika and Malawi are due to convergence, Molecular Phylogenetics and Evolution 2(2):158–165, 1993 г. Также см.: Williams, A., Taking firm hold of an illusion, J. Creation 20(3):45–49, 2006 г.; creation.com/Falk.
Edwards, C.J. и др., Ancient hybridization and an Irish origin for the modern polar bear matriline, Current Biology 21:1251–1258, 2011 г.
Sarfati, J., Design in living organisms (motors: ATP synthase), J. Creation 12(1):3–5, 1998 г.; creation.com/motor и Thomas, B., ATP synthase, Creation 31(4):21–23, 2009 г.; creation.com/ atp-synthase.
См. описание и анимацию в Smith, C., Fantastic voyage, Creation 30(1): 20–23, 2007 г.; creation.com/fantastic-voyage.
Christen, B. и др., The essential genome of a bacterium, Molecular Systems Biology 7:528, 2011 г. Жизненно важная часть генома бактерии Caulobacter crescentus, в разрешении 8 п.н., составила 480 открытых рамок считывания (белок-кодирующих генов), 402 регуляторных последовательности и 130 некодирующих элементов, в том числе 90 межгенных сегментов с неизвестной функцией; в общей сложности 492 941 пара оснований. Интересно, что из 320 жизненно необходимых белков Caulobacter, которые являются общими с E. coli (кишечной палочкой), более трети не являются незаменимыми для E. coli. Тем не менее, кишечная палочка имеет свои жизненно необходимые белки, которые отсутствуют у Caulobacter либо не являются незаменимыми для Caulobacter. Авторы пришли к выводу, что незаменимая часть генома бактерии зависит от взаимодействия всех компонентов конкретной бактерии. Например, АТФ-синтаза является незаменимой для Caulobacter, но не для E. coli, поскольку последняя может производить АТФ путём ферментации.
IAEA и FAO совместно публикуют вестник Plant Mutation Reports дважды в год.
Например: Catchpoole, D., Morning glory’s designer label clothing, Creation 29(1):49–51, 2006 г.; creation.com/morning-glorys-designer-label-clothing.
Mira A., Ochman H. и Moran N.A., Deletional bias and the evolution of bacterial genomes, Trends in Genetics 17(10):589–596, 2001 г.
Batten, D., Bacteria ‘evolving in the lab’? Июнь 2008 г.; creation.com/lenski.
Blount, Z.D. и др., Genomic analysis of a key innovation in an experimental Escherichia coli population, Nature 489:513–518, 2012 г.
См. обзор Batten, D., Clarity and confusion–A review of The Edge of Evolution, J. Creation 22(1): 28– 33, 2008 г.; creation.com/edge-evolution. Бехе также даёт комментарий по вопросу мутации E. coli, благодаря которой она получила способность усваивать цитрат: Behe, M., Rose-colored glasses: Lenski, citrate, and BioLogos, 13 ноября 2012 г.; www.evolutionnews.org/2012/11/rose- colored_gl066361.html.
omim.org/statistics/geneMap.
Haldane, J.B.S., The cost of natural selection, J. Genetics 55:511–524, 1957; Maynard Smith, J., The Theory of Evolution, Penguin Books, Хармондсворт, Великобритания, стр. 239, 1958 г.
Sanford, J. interviewed by Batten, D., Plant geneticist: «Darwinian evolution is impossible», Creation 30(4):45–47, 2008 г.; creation.com/sanford.
Доктор Сэнфорд представил этот анализ в своей книге «Генетическая энтропия и загадка генома» (Genetic Entropy and the Mystery of the Genome), FMS Publications; 3-е изд., март 2008 г. (доступно через creation.com). См. обзор первого издания: Truman, R., From ape to man via genetic meltdown: a theory in crisis–A review of Genetic Entropy & The Mystery of the Genome by John C. Sanford, J. Creation 21(1):43–47, 2007; creation.com/sanfordreview.
Sanford, J. interviewed by Batten, D., Plant geneticist: «Darwinian evolution is impossible», Creation 30(4):45–47, 2008 г.; creation.com/sanford.
См. Sanford, J., Critic ignores reality of Genetic Entropy, март 2013 г.; creation.com/genetic-entropy.
См. Sanford, J., Critic ignores reality of Genetic Entropy, март 2013 г.; creation.com/genetic-entropy.
Mendel’s Accountant; mendelsaccountant.info; Sanford, J. и др., Mendel’s Accountant, SCPE 8(2): 147–165, 2007; www.scpe.org/index.php/scpe/article/view/407/77.
Wieland, C., Blind fish, island immigrants and hairy babies, Creation 23(1):46–49, 2000 г.; creation.com/ blind-island.
Sarfati, J., Christopher Hitchens – blind to salamander reality, июль 2008 г.; creation.com/hitchens.
Wieland, C., Beetle bloopers, Creation 19(3):30, 2003 г.; creation.com/beetle-bloopers.
Wieland, C., Superbugs not super after all, Creation 20(1):10–13, 1997 г.; creation.com/superbugs.
Wieland, C., Rapid tomcod ‘evolution by pollution’? Февраль 2011 г.; creation.com/tomcod.
Catchpoole, D., The Stickleback: Evidence of evolution? Сентябрь 2009 г.; creation.com/stickleback.
Pennisi, E., Evolutionary biology: Changing a fish’s bony armor in the wink of a gene, Science 304(5678):1736–1739, 2004 г.
Coyne, J.A., Switching on evolution–how does evo-devo explain the huge diversity of life on Earth? Nature 435(7046):1029–1030, 2005 г.
Цит. по Catchpoole, ссылка 62.
Catchpoole, ссылка 62.
Lightner, J., interviewed by Batten, D., Getting it right, Creation 32(3):40–43, 2010 г; creation.com/creationist-veterinarian.
Lightner, J., Gain-of-function mutations, J. Creation 19(3):7–8, 2005 г.; creation.com/gain-of-function.
См., например, Whitelaw, E., Sins of the fathers, and their fathers, European J. Human Genetics 14:131–132, 2006 г.; Pembrey, M.E. и др., Sex-specific, male-line transgenerational responses in humans, European J. Human Genetics 14:159–166, 2006 г.; Jimenez-Chillaron, J.C. и др., Intergenerational transmission of glucose intolerance and obesity by in utero undernutrition in mice, Diabetes 58:460–468, 2009 г.
Morgan, H. и др., Epigenetic inheritance at the agouti locus in the mouse, Nature Genetics 23:314–318, 1999 г.
Carter, R., Darwin’s Lamarckism vindicated? Март 2011 г.; creation.com/epigenetics-and-darwin.
Haldane, J.B.S., The cost of natural selection, J. Genetics 55:511–524, 1957 г.
См. Tomkins, J. и Bergman, J., Genomic monkey business–estimates of nearly identical human- chimp DNA similarity re-evaluated using omitted data, J. Creation 26(1):94–100, 2012 г.; creation.com/chimp. Различие может составлять до 30%, как показал генетик Ричард Баггз: Buggs R., Chimpanzee? Октябрь 2008 г.; www.refdag.nl/chimpanzee_1_282611.
Batten, D., Haldane’s dilemma has not been solved, J. Creation 19(1):20–21, 2005 г.; creation.com/haldane. См. также ReMine, W., Cost theory and the cost of substitution–a clarification, J. Creation 19(1):113– 125, 2005 г.; creation.com/cost.
Catchpoole, D., Peacock tail tale failure, июнь 2008 г.; creation.com/peacock-tail-tale-failure.
Burgess, S., The beauty of the peacock tail and the problems with the theory of sexual selection, J.Creation 15(2):94–102, 2001 г.; creation.com/peacock.