
Происхождение жизни. Джонатан Сарфати, доктор философии (физическая химия) [Университет королевы Виктории в Веллингтоне, Новая Зеландия]
Университет королевы Виктории в Веллингтоне, Новая ЗеландияДоктор Сарфати широко известен в креационистских кругах, он опубликовал огромное количество статей и четыре сольные книги, а также ещё несколько книг в соавторстве. Одна из его работ, «Несостоятельность теории эволюции» (Refuting Evolution), является самой продаваемой креационистской книгой! Его работы отличает проницательная логика, возможно, благодаря его шахматному опыту (в 1988 г. он получил титул мастера ФИДЕ). Мы выбрали Джонатана для написания главы о третьей ахиллесовой пяте эволюции – происхождении жизни. Здесь он остроумно демонстрирует непреодолимое препятствие для возникновения первых живых организмов из неживых химических предшественников.
См. creation.com/jonathan-sarfati-russian
Введение
В предыдущей главе были рассмотрены проблемы, которые перед эволюцией ставит генетика. Но даже эти аргументы были весьма снисходительны к эволюции ввиду того, что они предполагают уже существующую самовоспроизводящуюся клетку с функционирующей генетической системой. Первая глава, о естественном отборе, также была снисходительной к эволюционной теории, поскольку эти аргументы применимы только к уже существующим сущностям, которые могут передавать далее любую отобранную информацию. Эта глава значительно поднимает ставки. Мы увидим, что потребовалось бы, чтобы эта первоначальная воспроизводящаяся система возникла сама по себе.
Вы узнаете, что происхождение первой жизни – это уязвимейшая ахиллесова пята для материалистов. Даже простейшие живые одноклеточные организмы являются чрезвычайно сложными и имеют многочисленные сложные механизмы, а также содержат инструкции по их построению, которые хранятся таким образом, что могут быть декодированы как для использования организмом, так и для передачи потомству. Простейшему гипотетическому самовоспроизводящемуся организму требуются множество белков и молекулярных механизмов, а также способ кодирования и хранения информации, необходимой для их выработки.169 Может ли такая клетка развиться из химических предшественников?
Биологическая информация в каждом известном на сегодняшний день самовоспроизводящемся организме на Земле хранится в ДНК, но эта информация не может быть прочитана без декодирующей системы. Однако инструкции по созданию такой декодирующей системы сами хранятся в ДНК. Можно ли разрешить эту проблему курицы и яйца? Кроме того, большинство из этих процессов используют энергию, получаемую из молекул АТФ, которые производит нанодвигатель под названием АТФ-синтаза. Но двигатель АТФ-синтаза не может быть образован без инструкций в ДНК, которые считываются декодирующей системой, использующей АТФ – получается тройственный круг, или, скажем, проблема яйца–личинки–кузнечика (что ещё хуже, чем проблема курицы и яйца). Есть ли решение у этой сложной головоломки, или это сигнал для нас, что происхождение жизни, какой мы её знаем, невозможно?
Некоторые предлагают решить эти проблемы, предполагая один тип молекулы, функционирующей и как система хранения/считывания информации, и как механизм для декодирования. Но насколько соответствует требованиям к первой жизни общепринятый на такую роль кандидат – РНК?
Кроме того, ненаправляемые химические реакции в гипотетическом первичном бульоне не будут иметь никаких шансов привести даже к этой минимальной сложности. Предполагаемые строительные блоки (аминокислоты) не смогут образовать достаточно длинные молекулы, необходимые для жизни; скорее наоборот – длинные молекулы будут распадаться. Многие из строительных блоков не образуются вообще либо слишком разбавлены и загрязнены, чтобы быть полезными. Кроме того, это будет смесь лево- и правовращающих форм вместо форм исключительно одного типа хиральности, задействованных в живых организмах. Разве эти факты не означают конец для всех сценариев происхождения жизни?
Наконец, исследования химической эволюции, как в пробирке, так и с помощью компьютерных симуляций, характеризуются недопустимым уровнем вмешательства разумного исследователя (которого не было у предполагаемого первичного бульона). Могут ли они считаться подтверждением происхождения жизни?
В довершение ко всему, ведущие исследователи соглашаются, что химическая эволюция признаётся не из-за фактов и доказательств, а из-за материалистической веры. Как тогда это может быть доказательством против утверждения Библии о том, что разумная Личность создала жизнь Своим актом божественной воли?
Эволюционные процессы не могут объяснить первую жизнь
Идея, что жизнь возникла путём химических процессов, изначально не укладывалась в эволюционную концепцию Дарвина, но большинство людей не знают об этом несоответствии. Сам Дарвин, похоже, имел трудности с происхождением жизни и тем, как вписать его в свою теорию. Он сделал несколько осторожных заявлений в печати и ещё несколько в личных письмах, но большая часть написанного им обнаруживает обычную попытку избежать выводов. Он знал, что старые идеи о самозарождении (таких созданий, как мыши или мухи) были опровергнуты Франческо Реди в 1668 году, однако затем вынужден был оспаривать мощные аргументы своего современника, учёного-креациониста Луи Пастера, который в прах развенчал самопроизвольное зарождение, даже микробов, в 1861 году,170 всего через два года после опубликования «Происхождения видов».
В заключительной главе первого издания «Происхождения видов» (1859 г.) Дарвин написал:
Я должен сделать вывод по аналогии, что, вероятно, все органические существа, которые когда-либо жили на этой земле, произошли от какого-то одного первозданного вида, в которого изначально вдохнули жизнь.171
Это не похоже на утверждение спонтанного зарождения жизни. Позже, в письме к другу в 1863 году, он раскаялся в своей формулировке, в то же время избегая самого ответа на вопрос:
Но я уже давно жалею, что в угоду общественному мнению использовал термин из Пятикнижия, относящийся к сотворению, под которым я на самом деле имел в виду «появились» посредством какого-то совершенно неизвестного процесса ... Это просто вздор – размышлять в настоящее время о происхождении жизни; можно с таким же успехом размышлять и о происхождении материи.172
Однако, как ни странно, он вставлял это заключительное предложение в каждое последующее издание «Происхождения видов»:
Есть величие в воззрении, что жизнь с её различными проявлениями Творец первоначально вдохнул в одну или несколько форм; и между тем как наша планета продолжала вращаться согласно неизменному закону тяготения, из такого простого начала эволюционировало и продолжает эволюционировать бесчисленное множество самых прекрасных и самых удивительных форм.
Понятно, что Дарвин верил в ненаправляемую эволюцию, но не ясно, верил ли он в химическую эволюцию. Затем, в 1871 году (в том же году, когда он опубликовал «О происхождении человека», в котором впервые чётко сформулировал эволюционную связь человека с низшими формами жизни), он углубился в контроверзу:
... если бы (и ох, какое большое если бы!) мы только могли себе представить, что в каком-то маленьком теплом водоёме, в котором присутствуют различные аммиачные и фосфорные соли, есть свет, тепло, электричество и т.д., химически сформировалось белковое [sic] соединение, готовое претерпеть ещё более сложные изменения ...173
Значит, даже Дарвин, отец современной эволюционной теории, колебался между двумя мнениями по наиболее важному вопросу: как возникла жизнь?
Знаменитый философ Энтони Флю (1923–2010) указал на проблему, непосредственно говоря об утверждениях ведущего атеиста-эволюциониста Ричарда Докинза174 (р. 1941):
Мне кажется, Ричард Докинз постоянно игнорирует тот факт, что сам Дарвин в четырнадцатой главе «Происхождения видов» отметил, что вся его аргументация начинается с существа, которое уже обладало репродуктивными способностями. По-настоящему всеобъемлющая теория эволюции должна дать объяснение тому, как эволюционировало это самое существо.
Сам Дарвин прекрасно понимал, что он не предложил такого объяснения. Теперь же мне кажется, что результаты более пятидесяти лет исследований ДНК приводят к новому и чрезвычайно мощному аргументу в пользу замысла.175
Это особенно примечательно, потому что доктор Флю до недавнего времени был известен как ведущий сторонник атеизма, но, к ужасу атеистического сообщества, незадолго до своей смерти отказался от этого убеждения.176 Одним из основных доводов, которые привели его к такому решению, была исключительная сложность даже простейших самовоспроизводящихся клеток.
Феодосий Добржанский (1900–1975), один из ведущих эволюционистов 20-го века и ярый материалист (несмотря на православное воспитание177), также понимал это. Говоря о происхождение жизни, он решительно отверг теоретиков, которые используют естественный отбор в качестве объяснения, потому что ему необходима уже существующая жизнь:
При чтении какой-либо другой литературы о происхождении жизни я боюсь, что не все авторы используют термин [естественный отбор] корректно. Естественный отбор – это дифференциальное воспроизведение, увековечение организма. Чтобы происходил естественный отбор, уже должны существовать самовоспроизводство или саморепликация и по крайней мере два различных самовоспроизводящихся объекта или сущности. … Я хотел бы призвать вас, пожалуйста, поймите, что нельзя использовать термин «естественный отбор» как вздумается. Добиологический естественный отбор – это противоречие терминов.178
Это различие стоит иметь в виду, продолжая чтение.
Химическая эволюция
Многие эволюционисты пытаются отвергнуть убедительные доказательства, приведённые в этой главе, утверждая, что происхождение жизни из неживых химических соединений не имеет ничего общего с эволюцией, и говоря, что правильным термином для этого процесса является абиогенез. Но их же собрат-эволюционист Горди Слэк порицает их за это:
Я думаю, неискренне утверждать, что происхождение жизни не имеет отношения к эволюции. Оно так же важно для неё, как и большой взрыв для физики или космологии. Эволюция должна быть способна объяснить, хотя бы теоретически, весь путь назад к самому первому организму, который мог воспроизводить себя посредством биологических или химических процессов. И чтобы в полной мере понять этот организм, мы просто обязаны знать, что было до него. На данный момент мы и близко не подошли к этому.179
Зоолог и физиолог (а также эволюционист) Джеральд Керкут (1927–2004) дал следующее определение общей теории эволюции (о которой также говорилось в главе 1): «теория, согласно которой все живые формы в мире возникли из единого источника, который в свою очередь произошёл из неорганической формы».180 Конечно, эта часть эволюции не может быть дарвиновской, как было показано выше. Её часто называют химической эволюцией. Например, сентябрьский номер журнала Scientific American за 1978 год был специально посвящён эволюции, и одна из главных статей называлась «Химическая эволюция и происхождение жизни». В ней говорилось:
Джон Холдейн, британский биохимик,181 кажется, был первым, кто понял, что восстановительная атмосфера, в которой не было свободного кислорода, была необходима для эволюции жизни из неживой органической материи.182
Давний стойкий приверженец теорий происхождения жизни, Сирил Поннамперума, в соавторстве опубликовал статью с таким же названием, при этом было указано, что он является сотрудником Лаборатории химической эволюции Департамента химии Мэрилендского университета.183 Кроме того, в 2011 году журнал Discover подготовил специальный выпуск «Эволюция: переосмысление истории жизни» (Evolution: Rethinking the Story of Life), и одна статья была, конечно же, о происхождении жизни.
Простая клетка?
Во времена Дарвина многие люди приняли на веру теорию спонтанного зарождения – что жизнь возникла из неживой материи. Поверить в это тогда было несколько легче, потому что структура клетки была практически неизвестна. В Германии Эрнст Геккель, известный как «бульдог Дарвина на континенте», утверждал, что клетка была «простым комочком белковых соединений углерода».184 Однако этому не было никакого оправдания, потому что уже ко временам Геккеля оптическая микроскопия чрезвычайно продвинулась, до уровня, который позволял различить многие субклеточные компоненты.185Как бы то ни было, научная революция в области молекулярной биологии, произошедшая за последние полстолетия, показывает, что клетке необходимы как высокое информационное содержание, так и способ передачи этой информации следующему поколению (воспроизведение).
Информация и химия
В предыдущей главе было показано, что ДНК хранит огромное количество закодированной информации. Атеист Ричард Докинз сам отметил:
Одна человеческая клетка обладает информационной ёмкостью, достаточной, чтобы три или четыре раза подряд сохранить все тридцать томов энциклопедии «Британика».186
Разница между живым и неживым заключается не в материи, а в информации. Живые организмы содержат потрясающие количества информации. Бо́льшая часть информации закодирована в ДНК ... 187
Докинз объясняет значительную часть этого естественным отбором, но, как было показано, такое пояснение не подойдёт для первой живой клетки. Поэтому все объяснения ограничены одной только химией. Однако, в самой химии строительных блоков ДНК нет ничего, что заставило бы их соединиться предопределённым образом, точно так же как силы взаимодействия между молекулами краски не могут заставить их соединяться в буквы и слова. Майкл Полани (1891–1976), бывший профессором физической химии в Университете Манчестера (Великобритания), который обратился к философии, поддержал это:
Точно так же, как порядок букв на печатной странице не связан с химией печатной страницы, так и последовательность оснований в молекуле ДНК не связана с химическими силами, действующими в молекуле ДНК. Именно эта физическая неопределённость последовательности обусловливает невероятность любой конкретной последовательности и тем самым позволяет ей иметь смысловое значение – смысловое значение, которое характеризуется математически детерминированным объёмом информации.188
Другими словами, информация в этой книге основана не на свойствах молекул чернил на бумаге (или пикселей на экране, если вы читаете её в электронном формате), а на том, как они упорядочены в буквы, слова, фразы, предложения и абзацы. Разлив чернила, вы не получите пьесы Шекспира! Также обратите внимание, что буквы не имеют смысла, если не понимать язык, в котором они используются. Например, слово «gift» по-английски означает «подарок», в то время как по-немецки – «яд». Неправильная языковая конвенция может означать неправильное сообщение. Один мой друг из Германии сказал мне, что когда он в первый раз попал в англоязычную страну, то подумал, что мы совсем спятили, потому что кругом были «магазины яда».
Информация, а не случай, является ключом к жизни. Информация не случайна (как слизь), и она не является закономерно-предсказуемой (как кристаллы, которые содержат мало информации). Ведущий эволюционный исследователь происхождения жизни 20-го века, Лесли Оргел (1927–2007), согласен с этим:
Живые существа отличаются упорядоченной сложностью. Кристаллы, такие как гранит, не могут быть отнесены к живым, потому что у них отсутствует сложность; смеси случайных полимеров не могут быть отнесены к живым, потому что у них отсутствует упорядоченность.189
Скорее информация – это неповторяющееся, непредсказуемое упоря- дочивание сигналов, которые могут быть прочитаны и поняты какой-то заранее спроектированной системой с заранее заданным набором правил хранения, поиска и интерпретации инструкций. Во всех современных формах информация создаётся разумным источником для передачи сообщения. И жизнь основана на информации. Как сказал физик и эволюционист Пол Дэвис (р. 1946):
Теперь мы знаем, что секрет жизни заключается не в химических ингредиентах как таковых, а в логической структуре и порядке соединения молекул. Подобно суперкомпьютеру, жизнь является системой обработки информации. … Именно программное обеспечение живой клетки, а не аппаратное, является настоящей загадкой.190
Но он ничего не предлагает для решения этой загадки. Вместо этого он продолжает: «Как глупые атомы смогли сами написать своё собственное программное обеспечение?» И отвечает: «Никто не знает», признавая, что «не существует ни одного известного физического закона, способного создать информацию из ничего».
Происхождение информации – это уязвимейшая ахиллесова пята эволюционной теории. Совершенно невозможно провести естественную аналогию с её появлением. Случайность – полная противоположность информации. То же самое и с точной закономерностью (как в кристаллах). Чтобы естественным образом возникла новая информация, не являющаяся ни случайной, ни бесконечно повторяющейся, в отсутствие ума или запрограммированного механизма, потребуется преодолеть такие огромные вероятностные барьеры, которые можно по праву назвать «невозможными».
Алекс Уильямс подробно объясняет это применительно к клеточным механизмам:
Полани указывал на подобные механизмам структуры, существующие в живых организмах ... Точно так же, как структуры и функции этих распространённых деталей механизмов не могут быть объяснены с точки зрения металла, из которого они сделаны, так и структура и функции параллельных компонент в жизни не могут сводиться к свойствам углерода, водорода, кислорода, азота, фосфора, серы и следовых микроэлементов, из которых они сделаны. Есть бесконечные примеры таких неупрощаемых структур в живых системах, но все они функционируют в рамках единого принципа, называемого «аутопоэзис» [«самосоздание»].191
Из той же статьи Вильямса:
Аутопоэзис является уникальным и удивительным свойством жизни, и в известной Вселенной нет ничего подобного ему. Он состоит из неупрощаемой иерархии структурированных уровней. Они включают: (I) компоненты с идеально чистым составом, (II) компоненты с высокоспецифичной структурой, (III), функционально интегрированные компоненты, (IV) всесторонне регулируемые информационно-управляемые процессы и (V) обратно-причинные метаинформационные стратегии для индивидуального и видового выживания ... Каждый уровень построен поверх более низкого уровня, но не может быть предсказан исходя из него. А между базовым уровнем (идеально чистый состав) и естественной средой существует непреодолимая пропасть.
Таким образом, конкретная форма биомолекул не может быть предсказана исходя из свойств водорода, кислорода и углерода. Кроме того, порядок букв ДНК не может быть предсказан исходя из химических свойств отдельных букв. На каждом уровне есть непреодолимая пропасть. Уильямс называет каждую такую пропасть невозможностью Полани, и он подробно рассмотрел множество подобных пропастей между живыми и неживыми системами. Эти пропасти представляют собой ещё одну ахиллесову пяту эволюционной мысли. Считать, что мир природы как-то нашёл способ обойти их, сродни вере в то, что все атомы водорода могут вдруг случайно выстроиться в прямую линию от одного конца Вселенной до другого. Ясно, что такого никогда не произойдёт, даже если концептуально это возможно. Это суть идеи, стоящей за невозможностью Полани.
Проблема «курицы и яйца» не разрешится
Примечательно, что система декодирования, которая переводит «язык» ДНК, сама закодирована в ДНК, и это настоящая проблема «курицы и яйца». Известный философ науки сэр Карл Поппер (1902–1994) отметил:
То, что делает происхождение жизни и генетического кода загадкой, ставящей нас в тупик, заключается в следующем: генетический код не имеет никакой биологической функции, если он не транслируется; то есть, если он не приводит к синтезу белков, структура которых определена кодом. Но ... механизмы, с помощью которых клетка ... транслирует код, состоят из по меньшей мере пятидесяти макромолекулярных компонентов, которые сами закодированы в ДНК. Таким образом, код может быть транслирован только с помощью определённых продуктов его трансляции.
Это создаёт замкнутый круг; похоже, действительно порочный круг, возникающий при любой попытке сформировать модель или теорию происхождения генетического кода.
Таким образом, мы можем столкнуться с возможностью того, что происхождение жизни (как и происхождение физики) станет непреодолимым барьером для науки и поставит крест на всех попытках свести биологию к химии и физике.192
Хотя Поппер написал это четыре десятилетия тому назад, происхождение генетического кода остаётся всё такой же загадкой, как и прежде. Недавно, в 2009 году, Докинз признал:
«Уловка-22» [прим. пер.: порочный круг, неразрешимая дилемма (по назва- нию романа Дж. Хеллера (1961 г.), в котором главный герой пытается обойти содержащий непреодолимое противоречие пункт 22 устава американской военной базы)] c происхождением жизни состоит в следующем: ДНК может реплицироваться, но для этого нужны ферменты, катализирующие этот процесс. Белки могут катализировать образование ДНК, но им нужна ДНК, определяющая правильную последовательность аминокислот.193
Действительно, происхождение генетического кода – это замкнутый круг: белковые механизмы необходимы для чтения ДНК, но инструкции для построения этих белковых механизмов сами закодированы в ДНК. Кроме того, они используют энергию, для чего необходимы молекулы АТФ, создаваемые нанодвигателем АТФ-синтаза. Но и он также закодирован в ДНК, которую декодируют механизмы, нуждающиеся в АТФ! Белки – это механизмы, а ДНК – репродуктивный материал, но и те, и другие необходимы одновременно, чтобы клетка вообще могла функционировать. И конечно, всё это было бы бесполезным, если бы не было никакой информации, которую можно воспроизвести.
Удивительные машины, участвующие в декодировании ДНК
Транскрипция и сморщивающий механизм
Даже первичное копирование корректной, кодирующей белок части ДНК в информационную РНК (или матричную РНК, мРНК) требует сложных механизмов. Они включают в себя фермент РНК-полимеразу, который состоит из четырёх белковых цепочек. Ещё один белок должен сообщать РНК-полимеразе, где начать чтение ДНК-кода. Комплекс ферментов затем двигается вдоль цепочки ДНК, добавляя соответствующие буквы РНК по одной, пока не остановится точно в нужном месте.
Ричард Эбрайт и его команда из Университета Ратджерс обнаружили ещё большую тонкость в процессе транскрипции.194 Именно эта транскрибированная мРНК затем транслируется в белки в сложных механизмах под названием рибосомы.
ДНК – двойная цепочка, и только одна из цепочек копируется, поэтому её нужно расплести перед копированием. Копирующий механизм, называющийся РНК-полимеразой, сначала закрепляется в точке начала гена (т.е. последовательности, кодирующей белок). Затем закреплённая РНК-полимераза затягивает в себя ДНК, что приводит к её «сморщиванию».195 В результате двойная спираль раскручивается, благодаря чему копия мРНК может быть сформирована с одной из нитей. Кроме того, раскручивание накапливает энергию, подобно скрученной резинке в игрушечном самолётике. И так же, как игрушечный самолёт, эта энергия в конце концов высвобождается, когда механизм открепляется от отправной точки и начинает двигаться вперёд. Также при этом раскрученная ДНК, которая выходит из задней части механизма, скручивается обратно (происходит «разморщивание»).
Трансляция и рибосома
Рибосома – жизненно важный механизм в клетке, который считывает информацию с мРНК и превращает её в белок. Даже в «простой» бактерии, такой как кишечная палочка (E. coli), которая живёт в кишечнике человека, рибосомы состоят из 50 различных белков и трёх различных рибосомных РНК (рРНК). В более сложных организмах, имеющих клеточные ядра (эукариотах), задействованы 73 различных белка и 4 рРНК. Один из экспертов заявил:
Рибосома, вместе со своими вспомогательными механизмами, – вероятно, самый сложный аппарат из когда-либо созданных. Все его компоненты являются активными и движущимися, а сам он – экологически чистый, поскольку производит только гуанозиндифосфат (ГДФ) и фосфат.196
Рибосомы также обеспечивают линейное выстраивание цепочки белка. Вне механизма растущая пептидная цепочка будет легко образовывать нежелательные боковые ответвления, поскольку боковые группы вступают в реакцию друг с другом (например, аминокислоты аспартат и глутамат имеют –СООН ветвь, которая может реагировать с –NH2 ветвью лизина или аргинина). При промышленном синтезе пептидов боковые группы должны быть блокированы защитными группами, а затем, когда синтез закончен, разблокированы удалением этих групп. Но в предполагаемом первичном бульоне не было химиков-органиков, которые могли бы сделать это в нужное время. Таким образом, если принимать во внимание это дополни- тельное ограничение реальной химии, то натуралистическое происхождение жизни представляется ещё гораздо более сомнительным. Жизнь основана на молекулах (ДНК, РНК и белках), которые не производятся вне специально созданных химических систем (химических заводов, к которым в том числе относятся живые клетки).
Не существует жизни без рибосом. Действительно, невозможно даже мысленно представить жизнь без них, поскольку базовая химия предотвращает повторяемое образование белков без сложных систем управления процессами, которые рибосома обеспечивает во всём живом.
Трансферная РНК
Молекулы транспортной РНК (тРНК) являются жизненно важными адаптерами, по форме напоминающими четырёхлистный клевер и отвечающими за перенос отдельных аминокислот к месту, где они будут добавлены в новый белок. Они состоят из около 80 нуклеотидных букв, три из которых называются антикодоном. Антикодон сочетается с соответствующим трёхбуквенным кодоном на мРНК. Таким образом, тРНК добавляют нужные аминокислоты в нужном месте к растущей пептидной цепи.
Кроме того, каждая аминокислота должна быть активирована, чтобы преодолеть энергетический барьер, который в обычных условиях предотвращает связывание с соседней аминокислотой в растворе. Энергия для этого процесса поступает из АТФ (будет рассмотрено ниже). Затем специальный фермент аминоацил-тРНК-синтетаза (АРСаза) в два этапа связывает каждую аминокислоту c соответствующей ей тРНК. Необходимо как минимум 20 АРСаз – по крайней мере по одной для каждого типа аминокислот. Любой сбой здесь разрушит сообщение и, таким образом, весь смысл генетического кода.
Молекулы-адаптеры тРНК должны иметь совершенно правильную геометрию, чтобы 1) держать аминокислоты в положении, в котором они могут формировать пептидную цепочку, и 2) размещать антикодон в нужном месте на мРНК. Правильная геометрия адаптеров работает только в контексте рибосомы (либо они должны были эволюционировать вместе, что проблематично до невозможности, либо они были разработаны вместе блестящим Конструктором). Любая дорибосомная жизнь не требовала бы специфической геометрии, так откуда же взялась эта характерная геометрия? Кроме того, гипотетически эволюционирующие адаптеры, скорее всего, взаимодействовали как друг с другом, так и с мРНК.197
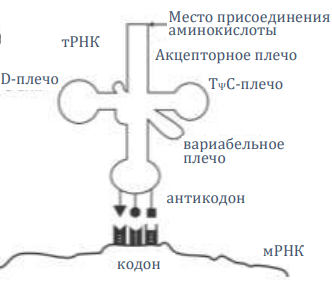
Как работает генетический код. Определенная группа из трех нуклеотидов на мРНК (кодон) взаимодействует с комплементарными нуклеотидами тРНК (антикодон). Поскольку тРНК с различными антикодонами несут различные аминокислоты, последовательность нуклеотидов мРНК определяет порядок аминокислот в итоговом белке. Каждая тРНК связывается с аминокислотой с помощью аминоацил-тРНК-синтетазы и молекулы АТФ (не показано). См. creation.com/gencode
Рибосомы создают в клетке контролируемую среду, со специфической геометрией, в которой может выполняться эта тонкая, но необходимая задача.
Кроме того, необходимо отсоединять тРНК-адаптеры, как только аминокислота присоединилась к концу растущей цепочки белка. Рибосома перемещается вдоль мРНК, как храповик, и энергия для отсоединения поступает из другой молекулы, хранящей энергию, – ГТФ (гуанозинтрифосфат), которая, в свою очередь, производится сложным и чётко интегрированным и регулируемым механизмом.198
Подводя итог: это невероятно сложная система. Как она могла возникнуть сама собой путём случайных химических реакций в первичном бульоне? Слишком много ограничений налагается на эволюционирующие системы, и слишком много возражений к процессу, возникающих при понимании основ химии.
Двойные сита – продвинутые химические механизмы
Аминоацил-тРНК-синтетазы (АРСазы) – это специальные ферменты (белковые механизмы), которые осуществляют удивительные химические процессы. Они даже могут отличить очень похожие химически аминокислоты (например, лейцин и изолейцин, которые особенно трудно отделить в лаборатории).199 Ведь, несмотря на то, что их различие небольшое, в важных биомолекулах оно может означать разницу между жизнью и смертью.
Однако АРСаза для изолейцина справляется с различением чрезвычайно хорошо: с долей ошибок всего 1 на 40000. Это достигается с помощью механизма двойного сита: одно сито отклоняет слишком большие аминокислоты, а другое – те, которые слишком малы.200
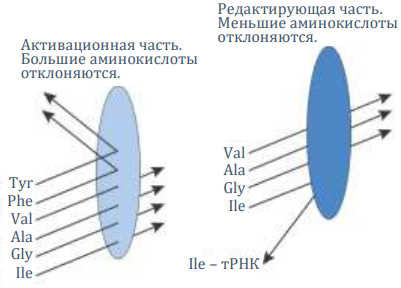
Механизм двойного сита изолейцил-тРНК-синтетазы. Гидролитическое редактирование снижает коэффициент ошибочной активации валина с ожидаемого значения между 1 из 10 и 1 из 100 до 1 из 40000. Fersht, A.R., Sieves in sequence, Science 280(5363)(ответ на статью Nureki и др., ссылка 32), 1998 г.
Шаперонины
Даже белковая цепочка, которая образуется в рибосоме, – это ещё не конечный продукт. Для выполнения своей функции в клетке белки, в том числе ферменты, о которых пойдёт речь ниже, должны быть правильно свёрнуты в свою специфическую трёхмерную форму. Окончательная конфигурация белка, произведённого согласно последовательности ДНК, определяется главным образом клеточными механизмами, называемыми шаперонинами, которые сами являются белками, по форме напоминающими бочку, и которые помогают другим белкам сворачиваться.201 Без шаперонинов важные белки могут сворачиваться неправильно, например в смертельные прионы. Это вероятная причина смертельного заболевания мозга Крейтцфельда–Якоба и губчатой энцефалопатии крупного рогатого скота (ГЭКРС), известной также как коровье бешенство.
Шаперонины также ограничивают эволюционный «прогресс», потому что отвергают неправильно свёрнутые белки, тем самым устанавливая ограничение на степень допустимых изменений. Для того чтобы новый белок эволюционировал, ему обязательно нужно было пройти ряд конформационных изменений, некоторые из которых индуцируют в структуре хаотические складки и изгибы. Но он также должен был как-то избежать или найти способ обойти фильтр шаперонина, специально предназначенный для избавления от неправильно свёрнутых белков. Эти механизмы, сворачивающие белки, широко распространены в живых организмах. Как могли высшие организмы эволюционировать после того, как ранние формы жизни начали использовать шаперонины? И как первые шаперонины правильно свернулись сами без уже существующих шаперонинов?202
Самый крошечный двигатель в мире – АТФ-синтаза
Ещё одним необходимым для жизни ферментом является АТФ-синтаза, которая производит энергетическую валюту организма – АТФ (аденозин- трифосфат). Энергия необходима для жизни, и все живые организмы используют АТФ в качестве энергетической валюты.203 Все живые существа, даже бактерии и археи, имеют двигатели АТФ-синтазы.204 Это, вероятно, делает АТФ-синтазу наиболее распространённым белковым комплексом на Земле.
Фактически тело человека с помощью многих триллионов этих двигателей ежедневно производит количество АТФ, примерно равное его собственному весу. АТФ потребляется очень быстро, поскольку обеспечивает энергией жизненно важные биохимические реакции, в том числе синтез ДНК и белков, мышечное сокращение, транспорт питательных веществ и передачу нервных импульсов. Организм без АТФ – как автомобиль без бензина, и некоторые яды (например, цианиды) действуют путём остановки производства АТФ.

Весь механизм АТФ-синтазы, состоящий из индивидуально произведённых белковых субъединиц, отмеченных буквами греческого алфавита. H+-ионы (протоны) переносятся через специальный туннель в АТФ-синтазе в направлении, указанном стрелкой. Это вызывает механическое движение, заставляя ось и основу вращаться вместе, подобно турбине. Почти 100% крутящего момента преобразуется в химическую энергию в форме молекул АТФ! Три молекулы АТФ производятся с помощью каждых 10 протонов.
Чтобы произвести молекулу АТФ, этот двигатель прижимает друг к другу два её компонента – АДФ (аденозиндифосфат) и фосфат – с достаточно высокой энергией. Затем он отбрасывает АТФ и готовится к принятию новых АДФ и фосфата. Этот мотор вращается со скоростью около 10000 оборотов в минуту, и при каждом обороте производятся три молекулы АТФ.
Фактически это два двигателя в одном. Верхняя часть (так называемая F1-АТФ-аза) состоит из трёх частей, каждая из которых – это АТФ-фабрика. Нижняя часть, FО,205 напрямую питается от положительно заряженного электрического тока (поток протонов), в отличие от отрицательно заряженного (поток электронов) тока, которым питаются разработанные человеком двигатели.206 Однако позже был обнаружен ещё один нюанс, связанный с тем, как эти двигатели соединены, а также с тем, как они собираются.207 А недавняя работа показывает, что это также самый эффективный двигатель в мире – фактически настолько эффективный, насколько позволяют законы физики. Исследователи пришли к выводу:
«Наши результаты свидетельствуют о 100% эффективности преобразования свободной энергии и плотном механохимическом сцеплении F1-АТФ-азы».208 Так что неудивительно, что даже сугубо секулярный журнал Nature назвал эти двигатели «настоящими двигателями создания».209
Ферменты
АТФ-синтаза и аминоацил-тРНК-синтетаза являются лишь двумя из многих видов белков, которые мы называем ферментами. Это биологические катализаторы, которые ускоряют жизненно важные химические реакции, при этом сами они не расходуются в ходе этих реакций. Жизни необходимы чрезвычайно эффективные ферменты, иначе выживание было бы невозможным: без них многие необходимые для жизни реакции были бы слишком медленными для её существования.210
Суперкатализаторы
В качестве хорошего примера чрезвычайно эффективного фермента Ричард Вулфенден в 1998 году продемонстрировал,211 что реакция, «“абсолютно необходимая” для создания строительных блоков ДНК и РНК, в воде протекала бы 78 миллионов лет»,212 но она была ускорена в 1018 раз213 ферментом.214 Чтобы оказывать такое воздействие, этот фермент должен иметь особую, чётко определённую структуру.215
В 2003 году Вулфенден открыл фосфатазу, которая катализирует гидролиз (расщепление) фосфатных связей и увеличивает скорость реакции в 1021 раз, что в тысячу раз больше, чем даже предыдущий фермент! Этот фермент позволяет реакции, играющей жизненно важную роль в клеточной сигнализации и регулировании, протекать за сотую долю секунды. Без фермента эта важнейшая реакция будет протекать триллион лет, то есть почти в сто раз больше, чем даже предполагаемый эволюционный возраст Вселенной (около 15 млрд лет)!216 Тем не менее эти ферменты, а также все другие процессы, перечисленные выше, должны были существовать в самой первой самовоспроизводящейся клетке, чтобы эта клетка могла выжить и передать ДНК потомству, а ДНК должна кодировать сами белки, необходимые для этих процессов!
Что из этого следует
Вулфенден заключил:
Без катализаторов не существовало бы никакой жизни вообще – от микроба до человека. Это заставляет задуматься, каким же образом действовал естественный отбор, чтобы произвести белок, который начал функционировать в качестве примитивного катализатора для такой чрезвычайно медленной реакции.217
Однако же, это любопытное слепое пятно. Как указывалось выше, естественный отбор не мог бы действовать до возникновения жизни, в то время как, по словам Вулфендена, жизнь не могла бы функционировать без этих ферментов, чрезвычайно ускоряющих важнейшие реакции. Происхождение жизни было изначально обречено на неудачу основными законами химии!
Простейшая возможная жизнь?
Более того, главной проблемой для возникновения жизни является именно информация, а не химия. Даже самая простая жизнь, которую только можно представить, должна иметь огромное информационное содержание. Бактерия Mycoplasma genitalium имеет наименьший известный геном среди всех известных организмов (вирусы не в счёт, потому что они полностью зависят от аппарата более сложных клеток для воспроизводства и сборки).218 Mycoplasma содержит 482 гена, состоящих из 580000 пар оснований.219 Конечно, эти гены функциональны только при наличии уже существующих транслирующих и реплицирующих механизмов, клеточной мембраны и т.д. Но у Mycoplasma genitalium нет собственной клеточной стенки, и она может выжить, только паразитируя на более сложных организмах (например, она живёт в клетках дыхательной системы и мочеполовых путей человека), обеспечивающих её многими питательными веществами, которые она не может производить для себя сама. По всей видимости, этот организм возник в результате потери генетической информации, что делает его зависимым от носителя.220
Микоплазма – это очень простые бактерии (хотя всё равно довольно сложные). Могло ли возникнуть что-то «проще»? В 1997 году Евгений Кунин, исследователь, занимающийся созданием искусственных биологических организмов, попытался подсчитать абсолютный минимум, необходимый для живой клетки. Он взял за основу своей работы микоплазму и попробовал подсчитать, без скольких генов даже эти простые клетки могут обойтись. Его команда пришла к результату 256 генов.221
Они сомневались, сможет ли такой гипотетический микроб долго существовать, ведь он едва ли способен ремонтировать повреждения своей ДНК, и теряет способность адаптироваться с оставшимся набором генов, усваивать сложные соединения, а также нуждается в обеспечении разнообразными органическими питательными веществами из окружающей среды.
Не удивительно, что в последующих исследованиях эта цифра была пересмотрена в сторону значительного увеличения. Этот новый гипотетический минимальный геном состоит из 387 генов, кодирующих белки, и 43 гена, кодирующих РНК.222
Статья за 2009 год в журнале New Scientist утверждает:
Нет никаких сомнений, что общий предок обладал ДНК, РНК и белками, универсальным генетическим кодом, рибосомами (фабриками по производству белков), АТФ и ферментом для производства АТФ, питаемым протонами. Также он должен был иметь ра́ звитые механизмы для считывания ДНК и преобразования генов в белки. Проще говоря, последний общий предок всего живого очень похож на современную клетку.223
В 2011 году биологи-эволюционисты, рассуждая об этом гипотетическом последнем универсальном общем предке, также пришли к выводу, что он не мог быть простым. Более того, он должен был включать «универсальные органеллы» для хранения высокоэнергетических соединений, называемых пирофосфатами; предыдущая догма утверждала, что у этого организма не было органелл.224 В одной статье говорится:
Как сообщают исследователи, новые данные показывают, что последний универсальный общий предок был всё-таки сложным организмом, со сложной структурой, в которой можно узнать клетку.225
Ни одна модель происхождения жизни и близко не подошла к этому абсолютному минимуму.
Могла ли такая сложность возникнуть случайно?
Естественный отбор не может действовать без самодостаточных и воспроизводящихся систем. Таким образом, он не может быть использован для объяснения этого минимального уровня сложности. Всё, что есть у эволюционистов, – это случай. И этот случай поддаётся достаточно простым расчётам вероятности.
Специалист по теории информации Хьюберт Йоки подсчитал, что даже при наличии пула чистых, активированных биологических аминокислот (намного более щедрое предложение, чем гипотетический «первичный бульон»), общий объём информации, которая могла быть произведена, даже с учётом миллиарда лет проб и ошибок, по временной шкале эволюционистов, будет всего лишь одним маленьким полипептидом длиной в 49 аминокислотных остатков.226 Речь идёт об одной восьмой длины (и соответственно, объёма информации) типичного белка, в то время как гипотетическая простая клетка требует по меньшей мере 387 белков (все предварительно закодированы в ДНК!), что позволило бы ей жить в очень специфической и инвариантной среде с постоянным обеспечением сложными питательными веществами и биомолекулами. Кроме того, расчёты Йоки щедро предполагают, что многие химические препятствия могут быть преодолены, а это является огромной натяжкой, как будет показано ниже.
Кроме того, можно вычислить вероятность получения последовательности ДНК для каждого из этих белков в случайном порядке. Конечно, есть некоторая свобода в последовательности для многих букв, но не возле активных центров. Тем не менее, даже эволюционные исследователи неявно признают, что некоторые последовательности критически важны. Они называют их «законсервированными» (т.е. последовательность была настолько значима, что естественный отбор сохранял её, устраняя вариации, которые появлялись в ходе эволюционной истории). Законсервированные целиком белки включают гистоны. Последние действуют как катушки, вокруг которых ДНК наматываются в хромосомах, убиквитин [прим. пер.: от англ. ubiquitous – вездесущий], который повсеместно используется в организмах, за исключением бактерий, и необходим для маркировки к уничтожению нежелательных белков,227 и калмодулин – повсеместно используемый белок, связывающий кальций, в котором почти все его 140–150 аминокислот «законсервированы».
Следующий расчёт будет очень щедр к эволюционистам. Мы представим, что только 10 аминокислот законсервированы в каждом ферменте и что есть какой-то механизм для присоединения аминокислоты в растворе для образования длинных цепочек (это слишком щедро, так как вода постоянно гидролизует пептидные связи в растворе):
20 аминокислот
387 белков для простейшей возможной жизни
10 законсервированных аминокислот в среднем
∴ вероятность равна 20–3870 = 10–3870∙log20 = 10–5035
Это один шанс на количество исследований, равное 1 с более чем 5000 нулями. Это было бы труднее, чем угадать правильный 5000-значный PIN-код с первой попытки!228 Однако, без такой совершенно случайной сборки жизнь невозможна. Это не вопрос медленного постепенного формирования дожизненных химических веществ, потому что речь идёт о самой простой форме жизни, по мнению тех, кто верит, что происхождение жизни было возможно в химическом бульоне.
Многие эволюционисты утверждают, что при наличии достаточного количества времени, возможно всё. Но действительно ли время является «героем» сюжета? Нет. Имеется:
1080 атомов во Вселенной
1012 взаимодействий атомов в секунду
1018 секунд с момента возникновения Вселенной, согласно дырявой теории большого взрыва
∴ только 10110 взаимодействий возможны.
Это огромное число, но по сравнению с количеством проб, необходимых, чтобы иметь реальные шансы получить правильную последовательность нуклеотидов, требуемых для кодирования нужных белков, которые необходимы для простейшей возможной формы жизни, оно абсурдно мало. Даже если бы все эти реакции происходили, вероятность была бы не более чем один шанс из 104925. Эти цифры настолько велики, что просто бессмысленны. Их суть в том, чтобы проиллюстрировать статистическую невозможность (речь идёт даже не о «невероятности») происхождения жизни из неживых химических веществ.
Знаменитый космогонист, сэр Фред Хойл (1915–2001), отказался от атеизма, признав абсурдно малые вероятности:
Вероятность формирования жизни из неживой материи – 1 к числу с 40 000 нулей после него… Этого достаточно, чтобы похоронить Дарвина и всю теорию эволюции. Не было никакого первичного бульона, ни на этой планете, ни на какой-либо другой, а если появление жизни было неслучайным, то оно должно было быть результатом целенаправленной работы разума.229
Самовоспроизводящиеся молекулы?230
Чтобы попытаться схватить за оба рога дилемму Поппера (представленную выше), как и приведённый анализ минимальной сложности жизни, некоторые эволюционисты предположили, что один тип молекулы мог выполнять как каталитическую, так и репродуктивную роль. Обычно выбирают между нуклеиновыми кислотами (РНК) и белками.
Однако, даже сейчас эволюционистам приходится признать, что РНК – действительно отвратительный катализатор, а белки – паршивые репликаторы. Ни один РНК-фермент и рядом не стоял по эффективности с белковыми ферментами, необходимыми для живых существ, подобных тем, которые анализировал доктор Вулфенден, и жизнь, какой мы её знаем, невозможна без такой эффективности. Бессмысленно также теоретизировать об альтернативных возможных формах жизни, потому что мы должны объяснить жизнь, какой мы её видим здесь, на планете Земля, и такая жизнь бросает вызов натуралистическим рассказам о происхождении. Эволюционисты признают, что рибозимы (РНК-ферменты, произведённые в лабораторных экспериментах) не являются эффективными ферментами; они никогда не могли достичь эффективности ферментов, необходимых для жизни. Даже Докинзу пришлось признать:
Дарвин в своём разделе о «тёплом маленьком водоёме» предположил, что ключевым событием в зарождении жизни могло быть спонтанное возникновение белка, но это предположение оказывается не столь перспективным, как большинство других идей Дарвина. ... Однако есть кое-что, в чём белки чрезвычайно плохи, и это Дарвин упустил из виду. Они совершенно безнадёжны в репликации. Они не могут создавать копии самих себя. Это означает, что ключевым шагом в зарождении жизни не могло быть спонтанное возникновение белка.231
Есть и другие огромные химические трудности с обеими идеями – «сперва РНК» и «сперва белки». Действительно, сторонники одной часто являются самыми резкими критиками другой. На самом деле обе стороны правы в своей критике, следовательно, жизнь не эволюционировала из химических веществ вообще!
Мир РНК
Очень популярная идея, что возникшая жизнь была основана на РНК, родилась в 1967 году, когда Карл Вёзе предположил, что РНК не только репродуктивна, но и могла также действовать как катализатор, таким образом выполняя обе роли.232 Томас Чек и Сидни Олтмен независимо друг от друга показали, что некоторые последовательности РНК обладают каталитическим действием. За «открытие каталитических свойств РНК» они получили Нобелевскую премию по химии в 1989 году.233
Обнаружение таких рибозимов позволило многим эволюционистам постулировать гипотезу о мире РНК. Они полагают, что первая жизнь состояла главным образом из молекул РНК, которые могли не только воспроизводить себя, но и осуществлять многие из функций, которые в настоящее время выполняют ферменты. Ричард Докинз иногда выступает сторонником этой идеи.234
Однако гипотезы мира РНК имеют ряд проблем:235
1. РНК – очень сложная молекула. Утверждение, что она могла возникнуть в первичном бульоне, – не более чем полёт фантазии. РНК даже менее стабильна, чем ДНК, а сама ДНК невероятно нестабильна (см. ниже). Даже составные компоненты РНК (нуклеотиды) сами по себе довольно сложные молекулы, которые не могли возникнуть в первичном бульоне. Химические процессы, необходимые для его синтеза в лаборатории, достаточно сложные.
2. Эксперименты с искровыми разрядами, такие как исследование Миллера–Юри, не приводят к появлению цитозина – одного из оснований РНК/ДНК. Сам цитозин, даже если бы он и мог возникнуть, является слишком нестабильным, чтобы накопиться в достаточном для использования количестве, даже в течение предполагаемого «длительного геологического времени», поскольку его период полураспада составляет 340 лет при 25 °C.236
3. Даже более простые строительные блоки РНК нестабильны вне клетки. Период полураспада рибозы составляет 44 года при pH = 7,0 и 0°C. При более высоких температурах всё ещё хуже (например, 73 минуты при 100°C).237 Основания РНК также очень быстро разрушаются в воде при 100°C, что является проблемой для теорий «тёплого водоёма», или гидротермальных теорий.238 Аденин и гуанин имеют периоды полураспада около года, урацил – около 12 лет, а цитозин – всего 19 дней.239
4. Подобно аминокислотам, о которых речь шла выше, нуклеотиды не способны спонтанно полимеризоваться; для этого они должны быть активированы. Более того, условия, оптимальные для усиления полимеризации, также способствуют разрушению рибозимов, и чтобы этот процесс осуществлялся, необходимы шаблоны.240 Ещё хуже то, что из-за повсеместного процесса гидролиза (разрушающего длинные цепочки биомолекул) вода обычно является врагом полимеризации.241
5. Химические реакции, в ходе которых создаются нуклеотиды, производят хиральные молекулы (оптические изомеры – зеркальные копии друг друга). Любая содержащая углерод молекула с четырьмя боковыми цепочками имеет «левовращающую» и «правовращающую» формы (см. рисунок), и любая реакция в первичном бульоне должна была произвести смесь таких форм в равных пропорциях (50/50).242 Чтобы сформировалась цепочка нуклеотидов, которая может свернуться спиралью (что необходимо для стабильности и реплицирования), все они должны
быть исключительно «односторонними» (или гомохиральными). Все пять нуклеотидных оснований (А, Ц, Г, Т и У) являются правовращающими, так же, как и все сахара, которые используются в остове ДНК. Интересно то, что все аминокислоты, используемые в живых организмах, – левовращающие. Из-за требования гомохиральности даже небольшая примесь молекул неправильной хиральности останавливает репликацию РНК не только в живых организмах, но и даже в искусственных системах.243
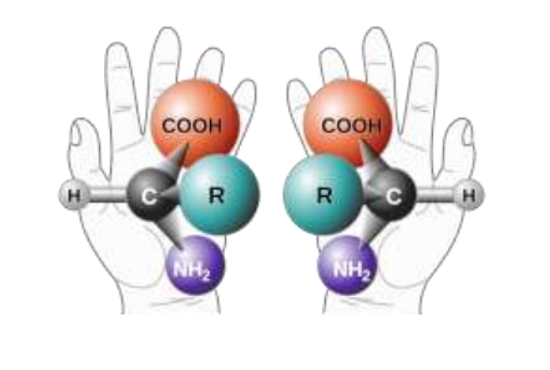
Два энантиомера обобщённой аминокислоты, где R – любая функциональная группа (кроме водорода H)
6. Даже если бы такие полимеры могли образоваться, что изначально должно было случиться без уже существующего шаблона, то они должны были бы обладать способностью к саморепликации (самовоспроизводству). Эта репликация должна была бы быть точной, в противном случае терялась бы любая информация, которую случайно удалось накопить. Даже точность в 96,7%, как в одном широко раструбленном случае,244 была бы совершенно недостаточной и привела бы к катастрофе ошибок. ДНК человека реплицируется с точностью примерно в одну ошибку на миллиард благодаря хорошо спроектированной, сложной системе исправления ошибок.
7. Если такие самореплицирующиеся молекулы РНК были основой первой формы жизни, они должны были бы выполнять все функции, необходимые для поддержания жизни организма. Однако фактически был обнаружен лишь ограниченный набор химических реакций, которые может выполнять РНК.
8. Помимо всего этого, как мог такой РНК-организм дать начало современным организмам, использующим белки-катализаторы, закоди рованные в репродуктивной ДНК? Это потребовало бы совершенно нового уровня устройств декодирования,245 а также невероятного переключения с информационной системы, основанной на РНК, на систему хранения и обработки информации, основанную на ДНК и белках.
9. Поскольку фосфаты – необходимый компонент нуклеиновых кислот, критически важно наличие свободных фосфатионов (PО43–), но учёные – сторонники мира РНК обычно используют их концентрацию, примерно в миллион раз превышающую ту, которая наблюдается в современных морях. В естественных условиях фосфат соединяется с избыточными ионами кальция (Ca2+) и магния (Mg2+), выпадая в нерастворимый осадок, при любых известных и гипотетических сценариях.246
Неудивительно, что один из ведущих исследователей моделей «мира РНК» Джеральд Джойс написал:
Наиболее вероятное предположение заключается в том, что жизнь не начиналась с РНК. … Проблема перехода к миру РНК, как и зарождения жизни в целом, полна неясностей и сталкивается с отсутствием экспериментальных данных.247
Другой исследователь химической эволюции, Роберт Шапиро, продемон- стрировав, что один из строительных блоков РНК не мог входить в состав первичного бульона, отметил:
Имеющиеся в настоящее время данные не подтверждают идею, что РНК или другой репликатор, использующий современный набор оснований РНК, присутствовали при возникновении жизни.
Отсутствие какого-либо правдоподобного стартового сценария происхождения жизни – это болезненная ахиллесова пята эволюционной теории. Дело не в том, что мы что-то упустили. Дело не в том, что, возможно, есть что-то, чего мы всё ещё не понимаем. Хотя это наверняка так, но всё, что мы уже знаем о физике, химии и вероятности, указывает против возможности происхождения жизни из неживых химических веществ.
Сперва белки
Более старая теория химической эволюции предполагала, что сперва появились белки. Она получила огромный пропагандистский импульс благодаря знаменитым экспериментам Миллера–Юри в 1950-х.
Эксперименты Миллера–Юри
Стэнли Миллер (1930–2007) был аспирантом Гарольда Юри (1893–1981), в 1934 году удостоенного Нобелевской премии по химии за открытие дейтерия (тяжёлого водорода).248 Миллер и Юри наполнили герметичный стеклянный аппарат восстановительными газами метаном, аммиаком и водородом (но исключили окисляющие газы, такие как кислород). Также аппарат имел колбу с кипящей водой для снабжения водой и обеспечения циркуляции газов внутри колбы с искровыми разрядами напряжением 60000 вольт, предназначенной для имитации молний. Газовая смесь проходила через конденсатор с водяным охлаждением, так что любые продукты реакции могли быть собраны путём попадания в водную ловушку внизу.
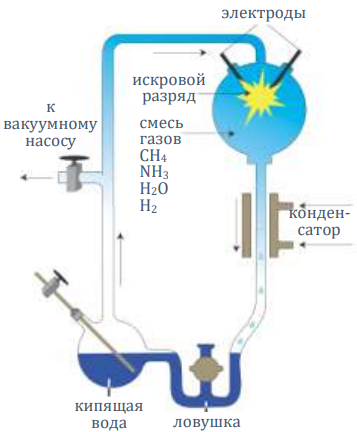
Спустя неделю в ловушке обнаружили красное пятно. Большей частью это была нерастворимая, токсичная, канцерогенная смесь – смола – обычный продукт органических реакций. В этой смоле было обнаружено небольшое количество аминокислот.249
Однако, это были, главным образом, самые простые аминокислоты глицин и аланин. Да и их количества были незначительными – 1,05% и 0,75% соответственно. Миллер признал: «Общий выход мал в сравнении с затраченной энергией».250 Тем не менее, нет никаких данных, что ситуация улучшится при наличии большего времени. Незначительные выходы простых продуктов – похоже, всё, что можно получить в результате этого моделирования.251
Забавно смотреть на сделанные предположения, неоднократно проведённые эксперименты и даже изготовленное хитроумное изобретение для получения незначительного выхода лишь некоторых аминокислот,252 которые не могут произвести ничего хотя бы отдалённо похожего на жизнь, и провозглашать, что «для возникновения жизни не был необходим разум»! В действительности Миллер и Юри обнаружили, что в результате случайных химических реакций возникают случайные химические вещества. Итак, произведённые химические вещества были также простыми. Случайная смесь простых молекул – противоположность тому, что необходимо для жизни.
Химия: смертельный враг спонтанного зарождения жизни
ДНК, информационная молекула клетки, на самом деле является очень нестабильной молекулой. В недавнем исследовании стабильности ДНК подсчитано, что, даже при хранении в кости, ДНК полностью распадётся за 22000 лет при 25°C, за 131000 лет при 15°С, за 882000 лет при 5°С и за 6,83 млн. лет при температуре –5°C.253 Другая статья сообщает:
«Существует распространённое мнение, что ДНК является «прочной как камень» – очень стабильной», – говорит Брандт Эйхман, адъюнкт-профессор биологических наук Университета Вандербильта, который руководил проектом. – «На самом деле ДНК обладает высокой реакционной способностью».254
Как уже упоминалось в главе 2, в среднем каждый день в клетке человека повреждается около одного миллиона оснований в ДНК. Эти повреждения вызваны сочетанием нормальной химической активности внутри клетки и воздействия радиации и токсинов, поступающих из окружающей среды, включая сигаретный дым, жареную еду и промышленные выбросы.255
Из-за такого высокого уровня ежедневных повреждений живые существа должны иметь сложную систему ремонта ДНК. Биолог Джеймс Шапиро из Университета Чикаго отмечает:
Все клетки, от бактериальных до человеческих, обладают поистине удивительным комплексом ремонтных систем, предназначенных для удаления случайных и стохастических источников мутации. Многоуровневая система корректирующих механизмов распознаёт и устраняет ошибки, которые неизбежно возникают в процессе репликации ДНК. Клетки защищают себя именно от тех видов случайных генетических изменений, которые, согласно общепринятой теории, являются источниками эволюционной изменчивости. Благодаря своим корректирующим и ремонтным системам живые клетки не являются пассивными жертвами случайных химических и физических воздействий. Они выделяют значительные ресурсы для подавления случайных генетических вариаций и имеют потенциал, чтобы установить уровень фоновой локализованной изменчивости путём регулирования активности своих ремонтных систем.
Конечно, гипотетический первичный бульон не содержал бы эти потрясающие ремонтные системы. Поэтому даже если бы ДНК удалось как-то сформироваться спонтанно, долго она бы не выжила.
РНК ещё более неустойчива, чем ДНК. Нестабильность РНК является основной причиной, по которой многие химические эволюционисты предпочитают сценарии «сперва белки». Однако белки также очень неустойчивы в долгосрочном масштабе и поэтому в любом случае будут разрушены. Они всё равно подчиняются второму закону термодинамики, и в конечном счёте будут разрушены из-за случайного движения атомов и фоновой радиации. Например, вода стремится разложить белки на составляющие их аминокислоты (гидролиз),256 также они подверглись бы деструктивным перекрёстным реакциям с другими химическими веществами257 в предполагаемом первичном бульоне.258 И так же, как в случае с нуклеотидами, описанном выше, они были бы в виде смеси лево- и правовращающих форм (рацемат), а не гомохиральными (только чисто лево- или чисто правовращающие формы), что необходимо для биологических ферментов.259
По самым последним оценкам, верхний предел периода устойчивости составляет 2,7 млн лет для коллагена и 110 млн. лет для костного белка остеокальцина при температуре замерзания воды (0°C). При всё ещё прохладной температуре в 10°C верхние пределы значительно ниже – 180000 лет для коллагена и 7,5 млн лет для остеокальцина. При 20°С максимальный период устойчивости ещё короче: 15000 и 580000 лет соответственно.260 Поскольку в целом скорость реакции увеличивается экспоненциально с повышением температуры (биологические реакции в среднем ускоряются вдвое при каждом повышении на 10°C),261 эта проблема становится неразрешимой для теории тёплого водоёма. Фактически неустойчивость белков – сильный аргумент против долгих веков. Мы находим их в костях динозавров, но если бы им на самом деле было более 65 млн лет, то не должно было остаться никаких обнаруживаемых белков.262 Кроме того, проблема для эволюционистов ещё более усложняется открытием ДНК в костях динозавров. Экспериментаторы исключили загрязнение, и ДНК достаточно хорошо сохранилась, чтобы образовать небольшие целые двойные спирали ДНК.263
Формирование белков из аминокислот
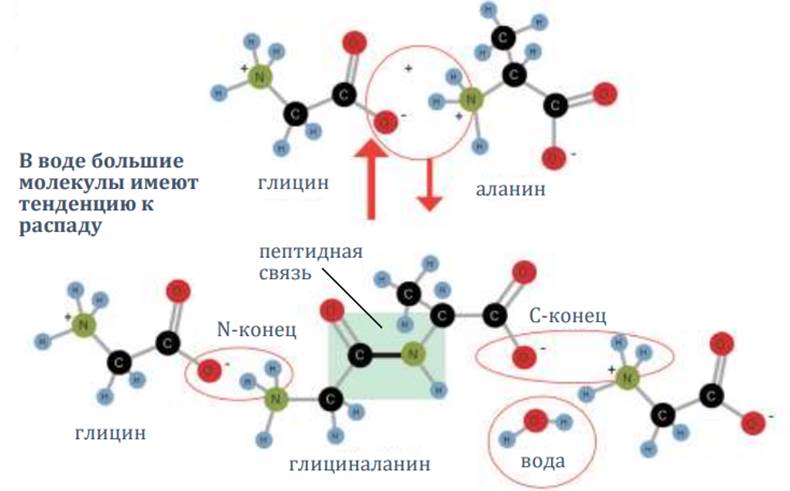
Чтобы сформировалась белковая цепочка, аминокислоты должны соединяться, создавая пептидные связи, с высвобождением молекулы воды. Но существует намного более сильная тенденция к обратному процессу. В воде она усиливается ещё больше.
Недопустимое вмешательство исследователей
Моделирование происхождения жизни призвано показать, что жизнь могла возникнуть благодаря времени и случаю. На самом деле даже те скудные полученные результаты возможны только благодаря планировке эксперимента. Обычная процедура такова: находят следы соединения А в эксперименте искрового разряда, а следы соединения B – в другом химическом эксперименте (иногда с взаимоисключающими условиями), а затем заявляют: «Смотрите, А и B могут быть получены в реалистичных условиях первобытной Земли». После берут чистые, гомохиральные, концентрированные А и B, синтезированные промышленными химическими компаниями, проводят реакцию между ними, чтобы получить следы более сложного соединения C.264 Затем по новостям трубят, что C могло образоваться в условиях первобытной Земли. Но это не значит, что разбавленные А и B способны реагировать таким образом, что могут образоваться вместе, или что они не будут вступать в реакцию с загрязняющими веществами D, E или F, которые также образовались в первых экспериментах. Короче говоря, моделирование эволюционистов демонстрирует недопустимый уровень разумного вмешательства.265
Большая часть эволюционистской пропаганды напоминает следующую гипотетическую теорию происхождения автомобиля:
Замысел – ненаучное объяснение, поэтому вместо него мы должны найти натуралистическое объяснение. Итак, эксперименты показали, что один из важных компонентов автомобиля – железо – может быть получен путём нагревания минералов естественного происхождения, таких как гематит, до температур, которые обнаруживаются в некоторых местах на Земле. Более того, можно показать, что железо может сформировать тонкие листы под давлением, которое, как известно, возникает в некоторых геологических формациях. ...
Если спонтанное происхождение автомобиля кажется надуманным, то обратите внимание, что даже простейшая самовоспроизводящаяся клетка имеет значительно более высокое информационное содержание, чем автомобиль, которому не требуется даже поддержание внутреннего гомеостаза, не говоря уже о воспроизведении.
Жизнь против «грязной» химии
Наши биомолекулы обретают свои свойства не благодаря своим составляющим самим по себе, а благодаря упорядочиванию этих составляющих, то есть информации. Такое упорядочивание не определяется физикой и химией этих составляющих, а наложено на них с более высокого уровня. Стоит вновь повторить то, что было сказано выше: свойства углерода, водорода, кислорода, азота, фосфора и серы не могут быть использованы для предсказания какой-либо конкретной макромолекулярной формы (существует бесконечное число возможностей). Скорее, макромолекулы были предназначены для выполнения конкретной функции с учётом свойств этих простых элементов.
Макромолекулы были сконструированы сверху вниз; они не эволюционировали снизу вверх. В живых организмах уже присутствующие механизмы накладывают эту упорядоченность (информацию) на компоненты, чтобы сделать больше копий, но сами эти механизмы возникли таким же образом, из предыдущего поколения, и так далее. Весь этот комплекс механизмов не возник из компонент, так же как белый шум не может создать симфонию.
Чем больше мы узнаём о биохимии, тем лучше понимаем, насколько она отличается от абиотической химии. Законы те же, но химия за пределами живой клетки – это всегда «грязная» химия, действующая с огромными объёмами молекул, в то время как биохимия – чистая помолекульная химия.266
Термин «грязная» ввёл исследователь химической эволюции и лауреат Нобелевской премии Кристиан де Дюв. Это всего лишь констатация факта, что химия в неживом мире одновременно оперирует с большим количеством молекул и неизбежно включает загрязняющие вещества.267 Палеонтолог, теистический эволюционист Саймон Конвей Моррис называл продукты типичных экспериментов по происхождению жизни «грязью», «слизью» и «гадостью»,268 вторя термину, который использовал исследователь химической эволюции Грэм Кэрнс-Смит, – «сильнозагрязнённая дрянь».269
Действительно, современная промышленная химия даже с помощью тщательно разработанных процессов обычно не может добиться чистоты выше 99,99%. Это в значительной степени потому, что она имеет дело с огромным количеством молекул одновременно. Но биохимия может быть намного эффективнее, потому что каждый фермент манипулирует одной молекулой за один раз. Затем за его продукт берётся следующий фермент, имеющий нужную форму и находящийся в нужном месте.
В конце концов, химия клетки точна, специализирована, управляема и функциональна, и если бы она не была таковой, то жизнь не могла бы существовать. Химия вне клетки противоположна тому, что необходимо для формирования жизни. Абиотическая химия – полная противоположность жизни. Как же она тогда могла привести к первой живой клетке?
Нереалистичные ловушки
Все источники энергии, которые производят биохимические вещества, уничтожают их несоизмеримо быстрее. Я уже упоминал гидролизирующее воздействие воды, которое приводит к разрушению пептидных связей гораздо быстрее, чем они образуются в растворе. Воздействие источников энергии также более разрушительное, нежели конструктивное. Например, на гипотетической первобытной Земле должны были присутствовать две формы ультрафиолетового (УФ) излучения. Одним из них является разрушительное (ионизирующее) коротковолновое ультрафиолетовое излучение. Другое – неразрушительное, с волной большей длины. Длинноволновое УФ-излучение в некоторых случаях может быть конструктивным. Например, оно может поглощаться и использоваться хлорофиллом.270 Однако, количество разрушительного УФ-излучения намного большее, а само оно более результативно, чем конструктивное УФ-излучение. Благодаря этим двум факторам деструктивные воздействия оказывают примерно в 104–105 раз более сильный эффект, чем конструктивные.271
В экспериментах Миллера–Юри использовались стратегически разработанные ловушки для изоляции биохимических веществ сразу после их формирования, так что разряды (или УФ-излучение в более поздних экспериментах) не уничтожали их. Без ловушек даже незначительные полученные количества не уцелели бы. Но это – нереалистичное моделирование первичного бульона, в котором отсутствует правдоподобный добиологический механизм для спасения аминокислоты от УФ путём быстрого их удаления из атмосферы, и даже от воздействия воды. Обратите внимание, что вы можете слегка загореть даже в пасмурный день, а иногда и под водой, поскольку УФ проникает даже сквозь десятки метров прозрачной жидкой воды.272 Так что ловушки были ещё одним примером неприемлемого уровня помех от разумных исследователей.
Химическая эволюция: доказательства или слепая вера?
Специалист по теории информации Хьюберт Йоки, не являющийся креационистом, сделал очень показательный комментарий более 30 лет назад:
Исследование происхождения жизни кажется уникальным в том плане, что вывод уже был авторитетно принят ... Остаётся найти сценарии, которые описывают подробные механизмы и процессы, посредством которых это произошло.273
Это важно иметь в виду, читая популярные изложения теории эволюции или отвечая тем, кто утверждает, что верящие в разумный замысел «предвзяты». Доктор Йоки закончил свою статью следующими словами:
Приходится сделать вывод, что, вопреки распространённому мнению, ещё не был предложен сценарий, описывающий происхождение жизни на Земле посредством случая и природных причин, который может быть признан на основе фактов, а не веры.274
Фонд происхождения жизни (Origin-of-Life Foundation) в настоящее время предлагает премию в 1 миллион долларов любому, кто предоставит химически правдоподобное натуралистическое объяснение происхождения генетического кода и жизни. На веб-странице говорится:
«Премия «Происхождение жизни»® (далее – «премия») будет вручена за предложение высоковероятного механизма спонтанного возникновения в природе генетических инструкций, достаточных для происхождения жизни. Для победы необходимо, чтобы объяснение согласовывалось с приведёнными ниже эмпирическими биохимическими, кинетическими и термодинамическими концепциями и было опубликовано в весьма уважаемом рецензируемом научном журнале (журналах).275
До сих пор не был вручён ни один приз, и чем больше мы знаем о минимальных требованиях к жизни, тем менее вероятным кажется, что будет найдено какое-нибудь хотя бы отдалённо правдоподобное материалистическое объяснение происхождения жизни. Проблема далека от решения, более, чем когда бы то ни было.276
Где доказательства?
Выше была критически проанализирована распространённая идея химической эволюции в первичном бульоне, но похоже, мало кто знает, что нет ни малейшего подтверждения тому, что он вообще когда-либо существовал. Такой бульон должен был быть источником необходимых азотсодержащих аминокислот и нуклеотидов. Если бы он существовал, эволюционные геологи должны были бы найти некоторые массивные богатые азотом отложения в (как они утверждают) очень древних породах. Тем не менее, азот практически отсутствует в том, что они называют древнейшими органическими материалами: его содержание составляет всего лишь около 0,015%. Два геохимика отмечают:
Если бы первичный бульон когда-либо существовал, то можно было бы ожидать найти хоть где-нибудь на этой планете массивные отложения, содержащие огромное количество различных азотистых органических соединений – кислот, пуринов, пиримидинов и им подобных; либо же мы должны были бы найти в сильно метаморфизованных отложениях огромное количество азотистых коксов. Фактически такие материалы нигде на Земле найдены не были.277
Жизнь из космоса?
Отчаяние теоретиков химической эволюции подчёркивает то, что некоторые исследователи утверждают, будто бы жизнь зародилась в космическом пространстве. Эта идея называется панспермией, от греческого πάν (пан, всё) и σπέρμα (сперма, семя), то есть семена жизни есть везде во Вселенной.
Классическое направление теории панспермии утверждает, что семена попали на Землю естественным путём, возможно с комет. Однако фактически это ничего не решает, а только переносит проблему в неизвестное место Вселенной. Химические и информационные проблемы остаются неизменными. Также, помимо невозможности выживания в течение миллионов лет дрейфования в холодном безвоздушном пространстве и при приближении к нашему Солнцу с его стерилизующими уровнями радиации, последние исследования показывают, что микробы не могут выдержать экстремальный нагрев, возникающий при трении с земной атмосферой, – нагрев, благодаря которому мы видим «падающие звезды».278
Более новая версия теории, которая называется направленной панспермией, утверждает, что жизнь на земле засеяли инопланетяне. В число её сторонников входят один из соавторов открытия двойной спирали ДНК Фрэнсис Крик (1916–2004), а также Лесли Оргел.279 Крика, в частности, к этому привело его разочарование в теориях химической эволюции:
Честный человек, вооружённый всеми знаниями, имеющимися у нас в настоящее время, вынужден признать, что в каком-то смысле зарождение жизни видится на данный момент практически чудом, настолько много условий должно было быть соблюдено, чтобы это произошло.
Каждый раз, когда я подготавливаю статью о происхождении жизни, я зарекаюсь в дальнейшем писать о нём, потому что в этом вопросе слишком много спекуляций, основанных на слишком малом количестве фактов.280
Опять же, это просто отодвигает вопрос на шаг назад; сторонники направленной панспермии должны верить, что эти гипотетические инопланетяне сами изначально возникли в результате химической эволюции.281 Интересно, как кто-то может критиковать сотворение как «ненаучное» из-за того, что оно постулирует ненаблюдаемого Творца, но при этом считать направленную панспермию «наукой», несмотря на ad hoc теорию ненаблюдаемых пришельцев. Отметим также, что панспермия является ещё одним примером аргумента «эволюции пробелов» и что направленная панспермия предполагает разумный замысел.
Выводы
Происхождение жизни из неживых химических веществ остаётся предметом слепой веры, а не науки, и являлось таковым со времён Дарвина. Основная проблема с теоретизированием по вопросу химической эволюции в том, что чаще всего теоретики рассматривают жизнь как набор химических соединений, а не как механизм обработки информации, и они никогда не дают ответ на вопрос: «Как же молекулярное оборудование смогло написать своё собственное программное обеспечение?»
Естественный отбор не может объяснить происхождение первой жизни. Чтобы обработать эту информацию, необходимы механизмы. Но эта информация включает в себя инструкции по созданию самих этих машин. Таким образом, проблем курицы и яйца предостаточно. Эти механизмы нуждаются в энергии, и она производится двигателем АТФ-синтаза. Но двигатель не может быть построен без инструкций и считывающих механизмов, закодированных в ДНК, а для сборки сборочных механизмов необходима АТФ. Это является проблемой яйца–личинки–кузнечика.
Другие ферменты необходимы для ускорения жизненно важных реакций, которые протекали бы миллионы лет без них. Однако, сами по себе ферменты оказывали бы разрушительное действие, например, изолированная АТФ-синтаза уничтожала бы АТФ, а не производила.
Белки не могут воспроизводить себя, и они бы распадались в естественных условиях. Длинные нуклеотиды и полисахариды также распадались бы (фактически они бы никогда и не сформировались) вследствие гидролиза. РНК является слабым ферментом и ещё более нестабильным, чем ДНК. Некоторые компоненты РНК так и не образовались в ходе экспериментов по «моделированию» химической эволюции. Другие получились разбавленными, загрязнёнными и/или неустойчивыми. Кроме того, они не были в необходимой требуемой для жизни гомохиральной форме.
Наконец, нет никаких подтверждений тому, что в «древнейших» породах происходили начальные и необходимые этапы добиологической химической эволюции.
В сущности, законы физики, химии и вероятности противоречат гипотезе абиотического происхождения жизни. В этом вопросе не библейские креационисты прибегают к аргументу «Бога пробелов»! Фактически мы утверждаем, что разумный Бог-Творец, который существует вне Вселенной и является причиной Вселенной, – единственный разумный вывод в свете того, что мы узнали за многие столетия экспериментальной науки.
Куда это ведёт?
Эта книга начинается с анализа главного дарвиновского двигателя эволюции – естественного отбора, и было показано, что он не оправдывает надежд. Затем мы обратились к генетике, о которой Дарвин был в совершенном неведении, хотя она необходима для полного понимания эволюции, и увидели, что эволюция и здесь терпит неудачу. Эти две главы составляют хороший куплет, поскольку рассматриваемые в них темы занимают центральное место в эволюционной теории. Настоящая глава о происхождении жизни может составить пару со следующей главой, написанной доктором Эмилем Сильвестру, о летописи окаменелостей.
Дарвину было необходимо, чтобы эти две области подтверждали его теорию, но поскольку наука была в зачаточном состоянии, то ему оставалось лишь надеяться, что они поддержат её в дальнейшем. Мы увидели, что Дарвин пытался избежать непосредственного обсуждения вопроса происхождения жизни, и не зря! Оказывается, что он также избегал подробного обсуждения летописи окаменелостей, поскольку знал, что она не подтверждает его идеи, исходя из имевшихся та тот момент данных. Если эволюция верна, то, во-первых, она должна быть возможна из химических и статистических соображений (а это не так), и во-вторых, мы должны увидеть подтверждения ей в палеонтологической летописи. Был ли Дарвин вправе предположить, что будущие открытия подтвердят медленные изменения на протяжении миллионов лет? Оказывается, что реальная летопись окаменелостей является ещё одной ахиллесовой пятой эволюции.
* * *
Без указания автора, Last universal common ancestor more complex than previously thought, ScienceDaily, 3 октября 2012 г.; sciencedaily.com.
См. Lamont, A., Louis Pasteur (1822–1895), Creation 14(1):16–19, 1991 г.; creation.com/louis-pasteur.
Darwin, C., On the Origin of Species, 1-е изд., John Murray, Лондон, 1859 г.
Письмо Джозефу Хукеру, 1863 г., Darwin, F. (ред.), из The Life and Letters of Charles Darwin, т. 3, стр. 18, John Murray, Лондон, 1887 г.; доступно через darwin-online.org.uk.
Darwin, F. (ред.), The Life and Letters of Charles Darwin, т. II, D. Appleton & Co., Нью-Йорк, стр. 202–203, 1911 г.
Детальные опровержения многих из самых популярных аргументов Докинза см. в книге Sarfati, J., The Greatest Hoax on Earth?, Creation Book Publishers, Паудер Спрингс, Джорджия, США, 2010 г., доступной через creation.com.
Flew, A., интервью Habermas, G., My pilgrimage from atheism to theism, Philosophia Christi, зима 2004 г.; biola.edu.
Flew, A., совместно с Varghese, R., There is a God, Harper Collins, New York, 2007 г. См. также рецензию на книгу: Cosner, L., J. Creation 22(3):21–24, 2008 г.; creation.com/flew.
Johnson, P.E., Response to Gould, Origins Research 15(1):10–11, 1993; www.arn.org. См. дополнительную информацию в O’Leary, D., Darwinist Theodosius Dobzhansky was not an orthodox Christian believer! post-darwinist.blogspot.com, 8 сентября 2006 г.
Dobzhansky, T.G., цит. по Schramm, G., Discussion of Synthesis of Nucleosides and Polynucleotides with Metaphoric Esters, в: Fox, S.W. (ред.), The Origins of Prebiological Systems and of Their Molecular Matrices, стр. 309–310, 1963 г., Academic Press, Нью-Йорк, 1965 г.
Slack, G., What neo-creationists get right, The Scientist, 20 июня 2008 г.; the-scientist.com.
Kerkut, G.A., Implications of Evolution, Pergamon, Oxford, стр. 157, 1960 г.
Джон Бёрдон Сандерсон Хо́лдейн (1892–1964) известен прежде всего как популяционный математический генетик, но он также работал с ферментами в Кембридже и вывел одно из уравнений ферментативной кинетики (Бриггса–Холдейна) в 1925 году. Будучи также убеждённым атеистом и коммунистом, он выдвинул версию натуралистического происхождения жизни в The Origin of Life, см. Rationalist Annual, стр. 148, 1928 г.
Dickerson, R.E., Chemical evolution and the origin of life, Scientific American 239(3):62–102, сентябрь 1978 г.
Pleasant, L.G. и Ponnamperuma, C., Chemical evolution and the origin of life, Origins of Life and Evolution of Biospheres 10(1):69–85, 1980 г.
Haeckel, E., The History of Creation, в переводе профессора E. Ray Lankester, M.A., F.R.S, 3-е изд., т. 1, стр. 184, Kegan Paul, Trench & Co., Лондон, 1883 г. См. также Grigg, R., Ernst Haeckel: Evangelist for evolution and apostle of deceit, Creation 18(2):33–36, 1996 г.; creation.com/haeckel.
Bradbury, S., The Microscope Past and Present, Pergamon Press, 1968 г.; van Niekerk, E., Countering Revisionism, J. Creation 27(1): 78–84, 2013 г.; creation.com/haeckel-fraud-proven-part-2.
Dawkins, R., The Blind Watchmaker, W.W. Norton, New York, стр. 115, 1986 г.
Dawkins, R., The Greatest Show on Earth, Free Press, New York, стр. 405, 2009 г.
Polanyi, M., Life’s irreducible structure, Science 160:1308, 1968 г.
Orgel, L., The Origins of Life, John Wiley, New York, стр. 189, 1973 г.
Davies, P., Life force, New Scientist 163(2204):27–30, 1999 г.
Williams, A., Life’s irreducible structure, J. Creation 21(1):109–115, 2007 г.; creation.com/autopoiesis.
Popper, K.R., Scientific Reduction and the Essential Incompleteness of all Science, в Ayala, F. и Dobzhansky, T. (ред.), Studies in the Philosophy of Biology, University of California Press, Berkeley, CA, США, стр. 270, 1974 г.
Dawkins, R., ссылка 19, стр. 420.
Revyakin, A. и др., Abortive initiation and productive initiation by RNA Polymerase involve DNA scrunching, Science 314(5802):1139–1143, 2006 г.; Kapanidis, A.N. и др., Initial transcription by RNA polymerase proceeds through a DNA-scrunching mechanism, Science 314(5802):1144–1147, 2006 г.; см. также Nanotech tools yield DNA transcription breakthrough; physorg.com, 16 ноября 2006 г
Roberts, J.W., RNA Polymerase, a scrunching machine, Science 314(5802):1139–1143, 2006 г.
Garrett, R., Mechanics of the ribosome, Nature 400(6747):811–812, 1999 г.
Truman, R. и Borger, P., Genetic code optimisation: Part 1, J. Creation 21(2):90–100, 2007 г.; сreation.com/gencode.
Truman, R. и Borger, P., ссылка 29.
Sarfati, J., Decoding and editing designs: double-sieve enzymes, J. Creation 13(1):5–7, 1999 г.; creation.com/doublesieve; Karlson, P., (перевод Doering, C.H.), Introduction to Modern Biochemistry, 4-е изд., Academic Press, Лондон и Нью-Йорк, стр. 113, 145–146, 1975 г.
Nureki, O. и др., Enzyme structure with two catalytic sites for double-sieve selection of substrate, Science 280(5363):578–582, 1998 г.
Sarfati, J., DNA: marvellous message or mostly mess? Creation 25(2):26–31, март 2003 г.; См. анимацию процесса трансляции ДНК по адресу creation.com/message.
Aw, S.E., The Origin of Life: A critique of current scientific models, J. Creation 10(3):300–314, 1996 г.; creation.com/origin-of-life-critique.
Bergman, J., ATP: The perfect energy currency for the cell, Creation Res. Soc. Q. 36(1):2–10, 1999 г.; creationresearch.org.
Hiroyuki Noji и др., Direct observation of the rotation of F1-ATPase, Nature 386(6622):299–302, 1997 г.
Обратите внимание, что это подстрочная буква О, а не цифра ноль, в силу исторических причин: от «часть, подавляемая олигомицином». Антибиотик олигомицин специфически блокирует протонный канал на FO части бактериальной АТФ-синтазы, что приводит к смерти бактерии
Sarfati, J., Design in living organisms (motors), J. Creation 12(1):3–5, 1998 г.; creation.com/motor. См. также Thomas, B., ATP synthase, Creation 31(4):21–23, 2009 г.; creation.com/atp-synthase. Подробнее см. в книге Sarfati, J., By Design, глава 10, «Motors», доступной через creation.com
Davies, K.M. и др., Macromolecular organization of ATP synthase and complex I in whole mitochondria, Proc. Nat. Acad. Sci. USA 108(34):14121–14126, 2011 г
Toyabea, S. и др., Thermodynamic efficiency and mechanochemical coupling of F1-ATPase, Proc. Nat. Acad. Sci. USA 108(44):17951–17956, 2011 г.
Block, S.M., Real engines of creation, Nature 386(6622):217–219, 1997 г. (обзор статьи Hiroyuki Noji и др., ссылка 36).
Катализаторы влияют не на равновесие, а только на скорость, с которой оно достигается. Они действуют за счёт снижения энергии активации, то есть уменьшают энергию переходного состояния или промежуточных продуктов реакции. См. диаграмму и объяснение к ней в Wieland, C. и Sarfati, J., Dino proteins and blood vessels: are they a big deal? 9 мая 2009 г.; creation.com/dino- proteins
Miller, B.G. и др., Anatomy of a proficient enzyme, Proc. Nat. Acad. Sci. USA 97(5):2011–2016, 2000 г.
Подразумевается, что реакция за 78 млн лет пройдёт наполовину. Использование полупериода реакции является распространённым способом для обозначения скорости реакции в химии.
Цит. по Lang, L.H., Without enzyme catalyst, slowest known biological reaction takes 1 trillion years: study, UNC School of Medicine 262:30, 2003 г.; unc.edu. См. также Wolfenden, R. и Snider, M.J., The depth of chemical time and the power of enzymes as catalysts, Acc. Chem. Res. 34:938–994, 2001.
Этим ферментом является оротидин-5'-монофосфат-декарбоксилаза, обеспечивающий синтез уридин-5'-фосфата – необходимого предшественника РНК и ДНК, – путём декарбоксилирования оротидин-5'-монофосфата (ОМФ).
Более подробную информацию можно найти в Sarfati, J., World record enzymes, J. Creation 19(2):13– 14, 2005 г.; creation.com/enzymes; а также в книге Sarfati, J, By Design, гл. 11, доступной через creation.com.
Lad, C., Williams, N.H. и Wolfenden, R., The rate of hydrolysis of phosphomonoester dianions and the exceptional catalytic proficiencies of protein and inositol phosphatases, Proc. Nat. Acad. Sci. USA 100(10):5607–5610, 2003 г.
Lang, L.H., ссылка 45
Однако вирусы имеют мощный наномотор, скручивающий их ДНК для компактности; Sarfati, J., Virus has powerful mini-motor to pack up its DNA, J. Creation 22(1):15–16, 2008 г.; creation.com/virusmotor; Fuller, D.N. и др., Single phage T4 DNA packaging motors exhibit large force generation, high velocity, and dynamic variability, Proc. Nat. Acad. Sci. USA 104(43):16868–16873, 2007 г.
Fraser, C.M. и др., The minimal gene complement of Mycoplasma genitalium, Science 270(5235):397– 403, 1995 г.; см. также обзор Goffeau, A., Life with 482 genes, Science 270(5235):445–446. Другие статьи содержат иные цифры, но все они не сильно отличаются.
Wood, T.C., Genome decay in the Mycoplasmas, Impact 340, 2001 г.; icr.org.
Wells, W., Taking life to bits, New Scientist 155(2095):30–33, 1997 г.
Glass, J.I. и др., Essential genes of a minimal bacterium, Proc. Nat. Acad. Sci. USA 103(2):425–430, 2006 г.
Lane, N., Was our oldest ancestor a proton-powered rock? New Scientist 204 (2730):38–42, 2009 г.
Seufferheld, M. и др., Evolution of vacuolar proton pyrophosphatase domains and volutin granules: clues into the early evolutionary origin of the acidocalcisomes, Biology Direct 6:50, 2011 г.
University of Illinois at Urbana-Champaign, ссылка 1.
Yockey, H.P., A Calculation of the probability of spontaneous biogenesis by information theory, J. Theor. Biol. 67:377–398, 1977 г.
Truman, R., The ubiquitin protein: chance or design? J. Creation 19(3):116–127, 2005 г.; creation.com/ubiquitin. Аарон Чехановер, Аврам Гершко и Ирвин Роуз получили Нобелевскую премию по химии в 2004 году «за открытие роли убиквитина в деградации белков»; nobelprize.org
На самом деле при очень низкой вероятности успеха p, а также при d = 1/р, хорошее эмпирическое правило гласит: чтобы вероятность проведения по крайней мере одного успешного испытания составляла 95%, необходимо провести 3d испытаний. В этом примере p составляет около 10–5000, то есть d = 105000, так что нам потребуется около 3 × 105000 испытаний для 95% вероятности получить все необходимые ферменты. Личная переписка с доктором Джимом Дэвидсоном (Северная Каролина), 2012 г.
Цит. по Major, E.L., Big enough to bury Darwin, Guardian (UK) education supplement, 23 августа 2001 г.; creation.com/hoyle-origin-of-life. См. также Demme, G. и Sarfati, J., Big-bang critic dies, J. Creation 15(3):6–7, 2001 г.; creation.com/hoyle.
Из Sarfati, J., Self-replicating enzymes? J. Creation 11(1):4–6, 1997 г.; creation.com/replicating.
Dawkins, R., ссылка 19, стр. 419–420
Woese, C., The Genetic Code, Harper and Row, New York, 1967 г
Press Release: The 1989 Nobel Prize in Chemistry, 12 октября 1989 г.; nobelprize.org
Dawkins, R., ссылка 18, стр. 421
См. также Mills, G.C. и Kenyon, D.H., The RNA world: a critique, Origins and Design 17(1): 9–16, 1996 г.; www.arn.org.
Shapiro, R., Prebiotic cytosine synthesis, Proc. Nat. Acad. Sci. USA 96(8):4396–4401, 1999 г.
Larralde, R., Robertson, M.P. и Miller, S. L., Rates of decomposition of ribose and other sugars: implications for chemical evolution, Proc. Nat. Acad. Sci. USA 92:8158–8160, 1995 г.
Sarfati, J., Hydrothermal origin of life?, J. Creation 13(2):5–6, 1999 г.; creation.com/hydrothermal.
Levy, M. и Miller, S. L., The stability of the RNA bases, Proc. Nat. Acad. Sci. USA 95(14):7933–7938, 1998 г.
Johnston W.K. и др., RNA-catalyzed RNA polymerization, Science 292(5520):131925, 2001 г.
Sarfati, J., Origin of life: the polymerization problem, J. Creation 12(3):281–284, 1998 г; creation.com/polymer.
Sarfati, J., Origin of life: the chirality problem, J. Creation 12(3):263–266, 1998 г.; creation.com/chirality.
Joyce, G.F. и др., Chiral selection in poly(C)-directed synthesis of oligo(G), Nature 310:602–604, 1984 г.
Johnston и соавторы (ссылка 72), признают, что точность копирования их рибозим является «всё ещё ниже, чем уровень точности ≥0.996, наблюдаемый у вирусных полимераз, которые реплицируют РНК с использованием РНК-шаблонов, и намного ниже, чем у полимераз, которые реплицируют ДНК».
Больший перечень химических проблем можно найти в Cairns-Smith, A.G., Genetic Takeover: And the Mineral Origins of Life, Cambridge University Press, 1982 г.; см. выдержку на creation.com/rna
Schirber, M., A salt-free primordial soup? Astrobiology Magazine, 19 января 2012 г.; astrobio.net.
Joyce, G.F., RNA evolution and the origins of life, Nature 338:217–224, 1989 г.
Биография Гарольда К. Юри на nobelprize.org.
Miller, S. L., A production of amino acids under possible primitive earth conditions, Science 117:528– 529, стр. 528, 1953 г.; Miller, S. L., Production of some organic compounds under possible primitive earth conditions, J. Amer. Chem. Soc. 77:2351–2361, 1955 г.
Йоки утверждал в Nature (415(6874):833, 2002 г.), что Стэнли Миллер не был первым. Прежде были эксперименты Уолтера Лёба (1913), Оскара Бодиша (1913), Эдварда Бейли (1922) и Гарольда Юри (1928, 1929). Йоки предположил, что Миллер просто дополнил эти предыдущие эксперименты современными методами разделения и обнаружения, такими как двумерная бумажная хроматография. Кстати, значение этих методов было подчёркнуто моим преподавателем органической химии. Ответ Джеффри Бада и Антониа Ласкано (Nature 416(6880):475, 2002 г.) отстаивал важность экспериментов Миллера для химической эволюции, в то время как Лёб не проявил к этому никакого интереса
Bergman, J., Why the Miller–Urey research argues against abiogenesis, J. Creation 18(2):74–84, 2002 г.; creation.com/urey
Cм. Sarfati, J., The Miller–Urey experiment revisited, creation.com/miller3-russian, 15 марта 2015 г
Allentoft, M.E. и др., The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils, Proc. Royal Society B 279(1748):4724–4733, 2012 г.
Newly discovered DNA repair mechanism, Science News; sciencedaily.com, 5 октября 2010 г.
Shapiro, J.A., A third way, Boston Review, стр. 2, февраль/март 1997 г.; Sarfati, J., New DNA repair enzyme discovered, creation.com/dna-repair, 13 января 2010 г
Sarfati, J., ссылка 74.
Например, аминогруппа (–NH2) в аминокислоте легко реагирует с карбонильной группой (O=C<) в сахаре, высвобождая молекулу воды (Н2O) с образованием имина (HN=C<), который для жизни бесполезен. См. Bergman, ссылка 83
Если такой бульон произвёл все необходимые для жизни азотистые соединения, то почему не осталось никаких их следов в «самых древних» породах? См. Brooks, J. и Shaw, G., Origins and Development of Living Systems, Academic Press, London, UK and New York, 1973 г
Sarfati, J., ссылка 74.
Nielsen-Marsh, C., Biomolecules in fossil remains: Multidisciplinary approach to endurance, The Biochemist, стр. 12–14, июнь 2002 г. См. также Doyle, S., The real ‘Jurassic Park’, Creation 30(3):12–15, 2008 г.; creation.com/realjurassic, и Thomas, B., Original animal protein in fossils, Creation 35(1):14–16, 2013 г.; creation.com/ancient-protein
Они подчиняются простому уравнению Аррениуса скорости реакции k = А ехр(−Ea/RT), где k – константа скорости, А – не зависящая от температуры постоянная (часто называемая «частотный фактор»), ехр – экспоненциальная функция, Еа – энергия активации, R – универсальная газовая постоянная, а Т – абсолютная температура.
Sarfati, J., ссылка 6, стр. 204–208
Schweitzer, M.H. и др., Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules, Bone 52(1):414–423, 2013 г.; см. также Sarfati, J., DNA and bone cells found in dinosaur bone, J. Creation 27(1):10–12, 2013 г.; creation.com/dino-dna-russian
Эволюционист Кэрнс-Смит выдвинул те же возражения против типичных экспериментов, моделирующих «происхождение жизни», в работе по ссылке 77
Thaxton, C.B., Bradley, W.L. и Olsen, R.L., The Mystery of Life’s Origin, гл. 6, Philosophical Library Inc., Нью-Йорк, 1984 г
Williams, A., Life’s irreducible structure–Part 1: autopoiesis, J. Creation 21(2):116–122, 2007 г.; creation.com/autopoiesis
De Duve, C., Singularities: Landmarks on the Pathways of Life, Cambridge University Press, Кембридж, 2005 г. См. обзор Williams, A., Great minds on the origin of life, J. Creation 21(1):38–42, 2007 г.; creation.com/singularities
Conway Morris, S., Life’s Solution: Inevitable humans in a lonely universe, Cambridge University Press, гл. 3–4, 2003 г. См. обзор ReMine, W., Evidence for Message Theory, J. Creation 20(2):29–35, 2006 г.; creation.com/lifes-solution
Cairns-Smith, A.G., Genetic Takeover and the Mineral Origins of Life, Cambridge University Press, Нью-Йорк, 1982 г
Фотосинтез является ещё одним примером неупрощаемо сложной системы, накапливающей энергию четырёх фотонов, чтобы расщепить сильно связанную молекулу воды. См. Sarfati, J., Green power (photosynthesis): God’s solar power plants amaze chemists, J. Creation 19(1):14–15, 2005 г.; creation.com/greenpower
Hulett, H.R., Limitations on prebiological synthesis, J. Theor. Biol. 24:56–72, 1969 г
Средневолновой ультрафиолет (UVB, 315–280 нм) проникает на глубину 65 метров в прозрачных водах Антарктики, согласно Gieskes, W.C. и Kraay, G.W., Transmission of ultraviolet light in the Weddell Sea: Report on the first measurements made in Antarctic, Biomass Newsl. 12:12–14, 1990 г
Yockey, H.P., ссылка 58, стр. 379
Yockey, H.P., ссылка 58, стр. 396
The Origin-of-Life Prize; www.us.net/life.
Smith, Calvin, Who wants to be a millionaire, creation.com/lifeprize, 15 августа 2007 г.
Brooks, J. и Shaw, G., Origins and Development of Living Systems, Academic Press, Лондон и Нью-Йорк, 1973 г.
Meteorite experiment deals blow to bugs from space theory, Physorg.com, 25 сентября 2008 г.; Sarfati, J., Panspermia theory burned to a crisp: bacteria couldn’t survive on meteorite, 10 октября 2008 г.; creation.com/panspermia
Crick, F. и Orgel, L.E., Directed Panspermia, Icarus 19:341–346, 1973 г
Crick, Life Itself, Its Origin and Nature, стр. 88, 153, Simon and Schuster, 1981 г
См. также Bates, G., Designed by aliens? Creation 25(4):54–55, 2003 г.; creation.com/aliens.